欢迎访问《中国农学通报》,
中国农学通报 ›› 2023, Vol. 39 ›› Issue (11): 1-9.doi: 10.11924/j.issn.1000-6850.casb2022-0293
• 农学·农业基础科学 • 下一篇
张彩娟1,2( ), 周坤能1, 夏加发1, 王元垒1, 云鹏1, 马廷臣1, 吴德祥2, 李泽福1()
), 周坤能1, 夏加发1, 王元垒1, 云鹏1, 马廷臣1, 吴德祥2, 李泽福1()
收稿日期:2022-04-14
修回日期:2022-08-27
出版日期:2023-04-15
发布日期:2023-04-10
通讯作者:
李泽福,男,1965年出生,安徽霍邱人,研究员,博士,主要从事水稻遗传育种研究。通信地址:230031 安徽省合肥市庐阳区农科南路40号 安徽省农业科学院水稻所,E-mail:作者简介:张彩娟,女,1993年出生,安徽亳州人,博士在读,研究方向:作物遗传育种。通信地址:230031 安徽省合肥市庐阳区农科南路40号 安徽省农业科学院水稻所,E-mail:zhangcaijuan1993@163.com。
基金资助:
ZHANG Caijuan1,2(), ZHOU Kunneng1, XIA Jiafa1, WANG Yuanlei1, YUN Peng1, MA Tingchen1, WU Dexiang2, LI Zefu1()
Received:2022-04-14
Revised:2022-08-27
Online:2023-04-15
Published:2023-04-10
摘要:
为了制定合理的育种计划,对籼型水稻光温敏核不育系‘7-163S’特征特性及基因型进行解析。以‘7-163S’及其配制组合或品种为试验材料,对主要农艺性状进行分析,使用基因组测序和RiceNavi数据库解析‘7-163S’的基因型。结果表明,‘7-163S’农艺性状优异,可繁性好,育性起点温度低、中抗稻瘟病、抗白叶枯病。已育成4个品种或组合,均表现出生育期适中、优质、丰产性好等特点。基因型分析表明‘7-163S’含有稻瘟病、白叶枯病、褐飞虱等抗性等位基因,以及低垩白、高氮肥利用效率、耐冷等优异等位基因。然而,其缺乏广谱抗稻瘟病基因Pi2、Pi9和Pigm、广谱抗白叶枯病基因Xa23、香味基因fgr及镉低积累基因OsCd1和OsNRAMP5等。将基因型与表型结合,可为基因聚合增强‘7-163S’抗性和新品种选育提供理论支撑。
张彩娟, 周坤能, 夏加发, 王元垒, 云鹏, 马廷臣, 吴德祥, 李泽福. 优质籼型水稻光温敏核不育系‘7-163S’特征特性及其基因型解析[J]. 中国农学通报, 2023, 39(11): 1-9.
ZHANG Caijuan, ZHOU Kunneng, XIA Jiafa, WANG Yuanlei, YUN Peng, MA Tingchen, WU Dexiang, LI Zefu. Characteristics and Genotype Analysis of High-quality Indica Rice Photo-Thermo-Sensitive Genic Male Sterile Line ‘7-163S’[J]. Chinese Agricultural Science Bulletin, 2023, 39(11): 1-9.
| 目标基因 | 引物名称 | 引物序列(5'-3') | 目的片段大小/bp(有/无) | 参考文献 |
|---|---|---|---|---|
| Pia | Pia-F | CTTTTGAGCTTGATTGGTCTGC | 116/125 | [ |
| Pia-R | CTATTGCACCAGAGGGACCAG | |||
| Pi2 | AP22-F | GTGCATGAGTCCAGCTCAAA | 140/155 | [ |
| AP22-R | GTGTACTCCCATGGCTGCTC | |||
| Pi9 | PB9-1-F | TAGACTCCTTCCAAGTTTGACT | 179/174 | [ |
| PB9-1-R | TGTGATTTTCAGAATTTTCGT | |||
| Pigm | DG-3-F | CAGAGCAGTAACAAACCCTA | 750/1800 | [ |
| DG-3-R | TCCGCAAGATCAACATTC | |||
| fgr | FGR3-F | GTATACCCCATCAATGGAAAT | 117/125 | [ |
| FGR3-R | CTTAACCATAGGAGCAGCTGA | |||
| Pik | RGA4-F | GGAAAGCTGATATGTTGTCG | 1650/无带 | [ |
| RGA4-R | ACTCGGAGTCGGAGAGTCAG | |||
| Pid3 | Pid3-dCAPS-F | TACTACTCATGGAAGCTAGTTCTC | BamH I酶切,510+148/658 | [ |
| Pid3-dCAPS-R | ACGTCACAAATCATTCGCTC | |||
| chalk5 | C1F | CGGTTAGAACAGGATTGC | 高垩白C1F/C1R2扩出197 bp,C1F/NC1R2无带 低垩白C1F/C1R2无带,C1F/NC1R2扩出197 bp | [ |
| C1R2 | ATAAAACAACTCTGGGTCACG | |||
| NC1R2 | ATAAAACAACTCTGGGTCACA | |||
| tms5 | tms5-F | ATATTTGGCGCTCTATTCTT | 387/463 | [ |
| tms5-R | GGCCAAGTGTTATGATCACT | |||
| Bph15 | MS5-F | TTGTGGGTCCTCATCTCCTC | 200/小于200 | [ |
| MS5-R | TGACAACTTGTGCAAGATCAAA |
| 目标基因 | 引物名称 | 引物序列(5'-3') | 目的片段大小/bp(有/无) | 参考文献 |
|---|---|---|---|---|
| Pia | Pia-F | CTTTTGAGCTTGATTGGTCTGC | 116/125 | [ |
| Pia-R | CTATTGCACCAGAGGGACCAG | |||
| Pi2 | AP22-F | GTGCATGAGTCCAGCTCAAA | 140/155 | [ |
| AP22-R | GTGTACTCCCATGGCTGCTC | |||
| Pi9 | PB9-1-F | TAGACTCCTTCCAAGTTTGACT | 179/174 | [ |
| PB9-1-R | TGTGATTTTCAGAATTTTCGT | |||
| Pigm | DG-3-F | CAGAGCAGTAACAAACCCTA | 750/1800 | [ |
| DG-3-R | TCCGCAAGATCAACATTC | |||
| fgr | FGR3-F | GTATACCCCATCAATGGAAAT | 117/125 | [ |
| FGR3-R | CTTAACCATAGGAGCAGCTGA | |||
| Pik | RGA4-F | GGAAAGCTGATATGTTGTCG | 1650/无带 | [ |
| RGA4-R | ACTCGGAGTCGGAGAGTCAG | |||
| Pid3 | Pid3-dCAPS-F | TACTACTCATGGAAGCTAGTTCTC | BamH I酶切,510+148/658 | [ |
| Pid3-dCAPS-R | ACGTCACAAATCATTCGCTC | |||
| chalk5 | C1F | CGGTTAGAACAGGATTGC | 高垩白C1F/C1R2扩出197 bp,C1F/NC1R2无带 低垩白C1F/C1R2无带,C1F/NC1R2扩出197 bp | [ |
| C1R2 | ATAAAACAACTCTGGGTCACG | |||
| NC1R2 | ATAAAACAACTCTGGGTCACA | |||
| tms5 | tms5-F | ATATTTGGCGCTCTATTCTT | 387/463 | [ |
| tms5-R | GGCCAAGTGTTATGATCACT | |||
| Bph15 | MS5-F | TTGTGGGTCCTCATCTCCTC | 200/小于200 | [ |
| MS5-R | TGACAACTTGTGCAAGATCAAA |
| 名称 | 7-163S | |
|---|---|---|
| 加工品质 | 糙米率/% | 80.5± 0.3 |
| 精米率/% | 73.1 ±0.2 | |
| 整精米率/% | 68.6 ±0.6 | |
| 精米粒长/mm | 6.2 ±0.2 | |
| 精米长宽比 | 3.0 ±0.0 | |
| 外观品质 | 垩白米率/% | 11.0 ±2.0 |
| 垩白度/% | 0.6 ±0.2 | |
| 透明度 | 2.0 ±0.0 | |
| 理化性质 | 碱消值 | 6.2 ±0.1 |
| 直链淀粉含量/% | 16.1 ±0.1 | |
| 胶稠度/mm | 86.0 ±1.0 | |
| 蛋白质含量/% | 9.8 ±0.2 | |
| 米质等级 | 2 | |
| 抗性鉴定 | 稻瘟病 | 中抗稻瘟病(MR) |
| 白叶枯 | 抗白叶枯病(R) | |
| 名称 | 7-163S | |
|---|---|---|
| 加工品质 | 糙米率/% | 80.5± 0.3 |
| 精米率/% | 73.1 ±0.2 | |
| 整精米率/% | 68.6 ±0.6 | |
| 精米粒长/mm | 6.2 ±0.2 | |
| 精米长宽比 | 3.0 ±0.0 | |
| 外观品质 | 垩白米率/% | 11.0 ±2.0 |
| 垩白度/% | 0.6 ±0.2 | |
| 透明度 | 2.0 ±0.0 | |
| 理化性质 | 碱消值 | 6.2 ±0.1 |
| 直链淀粉含量/% | 16.1 ±0.1 | |
| 胶稠度/mm | 86.0 ±1.0 | |
| 蛋白质含量/% | 9.8 ±0.2 | |
| 米质等级 | 2 | |
| 抗性鉴定 | 稻瘟病 | 中抗稻瘟病(MR) |
| 白叶枯 | 抗白叶枯病(R) | |
| 材料 | 鉴定条件 (光照时间-温度) | 花粉观察日期 | 育性 | |
|---|---|---|---|---|
| 花粉败育度/% | 自交结实率/% | |||
| 7-163S | 14.5 h-23.5℃ | 8/10-19 | 99.19 | 0.41 |
| 14.5 h-24℃ | 8/6-12 | 99.36 | 0 | |
| 12.5 h-25.5℃ | 8/5-12 | 99.45 | 0 | |
| 14.5 h-自然 | 8/2-9 | 99.63 | 0 | |
| 培矮64S | 14.5 h-23.5℃ | 8/10-19 | 99.31 | 0.35 |
| 14.5 h-24℃ | 8/6-12 | 99.43 | 0 | |
| 12.5 h-25.5℃ | 8/5-12 | 99.46 | 0 | |
| 14.5 h-自然 | 8/2-9 | 99.55 | 0 | |
| 材料 | 鉴定条件 (光照时间-温度) | 花粉观察日期 | 育性 | |
|---|---|---|---|---|
| 花粉败育度/% | 自交结实率/% | |||
| 7-163S | 14.5 h-23.5℃ | 8/10-19 | 99.19 | 0.41 |
| 14.5 h-24℃ | 8/6-12 | 99.36 | 0 | |
| 12.5 h-25.5℃ | 8/5-12 | 99.45 | 0 | |
| 14.5 h-自然 | 8/2-9 | 99.63 | 0 | |
| 培矮64S | 14.5 h-23.5℃ | 8/10-19 | 99.31 | 0.35 |
| 14.5 h-24℃ | 8/6-12 | 99.43 | 0 | |
| 12.5 h-25.5℃ | 8/5-12 | 99.46 | 0 | |
| 14.5 h-自然 | 8/2-9 | 99.55 | 0 | |
| 名称 | 两优799 | 两优华363 | 两优华166 | 永两优206 |
|---|---|---|---|---|
| 产量/(kg/hm2) | 9178.6±238.9 | 9245.7±71.6 | 9571.1±83.3 | 9325.6±77.5 |
| 株高/cm | 125.6±0.6 | 121.0±1.9 | 118.3±0.5 | 122.5±1.7 |
| 穗长/cm | 23.8±0.2 | 23.5±0.1 | 24.4±0.9 | 23.1±0.8 |
| 每穗总粒数 | 193.6±9.9 | 204.9±10.0 | 211.9±9.1 | 189.0±7.0 |
| 每穗实粒数 | 157.2±6.2 | 172.6±7.5 | 161.4±0.5 | 151.0±8.0 |
| 结实率/% | 81.2±1.1 | 84.2±0.5 | 80.6±3.9 | 79.9±1.1 |
| 千粒重/g | 26.9±0.3 | 24.5±0.3 | 25.9±0.0 | 22.2±0.7 |
| 全生育期/d | 140.2±5.4 | 140.6±4.5 | 141.6±7.2 | 135.9±0.5 |
| 米质等级 | 2 | 1 | 3 | 2 |
| 产量与对照相比增减产/% | 6.0±1.4 | 7.1±0.3 | 7.5±0.2 | 1.5±3.3 |
| 名称 | 两优799 | 两优华363 | 两优华166 | 永两优206 |
|---|---|---|---|---|
| 产量/(kg/hm2) | 9178.6±238.9 | 9245.7±71.6 | 9571.1±83.3 | 9325.6±77.5 |
| 株高/cm | 125.6±0.6 | 121.0±1.9 | 118.3±0.5 | 122.5±1.7 |
| 穗长/cm | 23.8±0.2 | 23.5±0.1 | 24.4±0.9 | 23.1±0.8 |
| 每穗总粒数 | 193.6±9.9 | 204.9±10.0 | 211.9±9.1 | 189.0±7.0 |
| 每穗实粒数 | 157.2±6.2 | 172.6±7.5 | 161.4±0.5 | 151.0±8.0 |
| 结实率/% | 81.2±1.1 | 84.2±0.5 | 80.6±3.9 | 79.9±1.1 |
| 千粒重/g | 26.9±0.3 | 24.5±0.3 | 25.9±0.0 | 22.2±0.7 |
| 全生育期/d | 140.2±5.4 | 140.6±4.5 | 141.6±7.2 | 135.9±0.5 |
| 米质等级 | 2 | 1 | 3 | 2 |
| 产量与对照相比增减产/% | 6.0±1.4 | 7.1±0.3 | 7.5±0.2 | 1.5±3.3 |
| 基因名称 | 基因号 | 农艺性状 | 突变型与野生型相比的表型变化 | 7-163S类型 | 7-163S表型 |
|---|---|---|---|---|---|
| NOG1 | LOC_Os01g54860 | 产量组成相关 | 每穗粒数增加 | 突变型 | 每穗粒数增加 |
| LAX1 | LOC_Os01g61480 | 产量组成相关 | 抗旱性提高、产量提高 | 突变型 | 抗旱性提高、产量提高 |
| DPL1 | LOC_Os01g15448 | 产量组成相关 | 杂交不育 | 野生型 | 杂交可育 |
| tms5 | LOC_Os02g12290 | 产量组成相关 | 温敏不育 | 突变型 | 温敏不育 |
| GW2 | LOC_Os02g14720 | 产量组成相关 | 籽粒变大,柱头不外露 | 野生型 | 籽粒变小 |
| OsGRF4 | LOC_Os02g47280 | 产量组成相关 | 氮肥吸收效率提高 | 野生型 | 氮肥吸收效率一般 |
| GNP1 | LOC_Os03g63970 | 产量组成相关 | 每穗粒数增加,株高增高 | 突变型 | 每穗粒数增加,株高增高 |
| NAL1 | LOC_Os04g52479 | 产量组成相关 | 增加叶宽、提高产量 | 突变型 | 增加叶宽、提高产量 |
| GW5 | LOC_Os05g09520 | 产量组成相关 | 粒宽减小 | 野生型 | 粒宽增加 |
| S5 | LOC_Os06g11010 | 产量组成相关 | 广亲和 | 野生型 | 籼粳杂交不亲和 |
| GW8 | LOC_Os08g41940 | 产量组成相关 | 粒宽增加,粒长减少 | 突变型 | 粒宽增加,粒长减少 |
| Rf4 | LOC_Os10g35240 | 产量组成相关 | 恢复野败型不育系育性 | 突变型 | 恢复野败型不育系育性 |
| LOX-3 | LOC_Os03g49260 | 产量组成相关 | 稻米储藏不产生腐败味 | 野生型 | 稻米储藏产生腐败味 |
| SKC1 | LOC_Os01g20160 | 非生物胁迫 | 耐盐性提高 | 野生型 | 耐盐性降低 |
| CYP72A31 | LOC_Os01g41800 | 非生物胁迫 | 除草剂耐受性增强 | 野生型 | 除草剂耐受性低 |
| NRAT1 | LOC_Os02g03900 | 非生物胁迫 | 铝胁迫敏感 | 野生型 | 耐铝胁迫 |
| OsHMA4 | LOC_Os02g10290 | 非生物胁迫 | 铜含量降低 | 野生型 | 铜含量高 |
| CAL1 | LOC_Os02g41904 | 非生物胁迫 | 减少镉积累 | 野生型 | 镉积累 |
| qLTG3-1 | LOC_Os03g01320 | 非生物胁迫 | 耐低温萌发能力提高 | 突变型 | 耐低温萌发能力提高 |
| OsCd1 | LOC_Os03g02380 | 非生物胁迫 | 镉富集增多 | 突变型 | 镉富集增多 |
| OsJAZ1 | LOC_Os04g55920 | 非生物胁迫 | 降低根长、根重 | 野生型 | 提高根长、根重 |
| OsHKT2;1 | LOC_Os06g48810 | 非生物胁迫 | 钾利用效率降低 | 野生型 | 钾利用效率提高 |
| OsHMA3 | LOC_Os07g12900 | 非生物胁迫 | 富集镉 | 野生型 | 不富集镉 |
| OsNRAMP5 | LOC_Os07g15370 | 非生物胁迫 | 富集镉 | 突变型 | 富集镉 |
| OsSAP16 | LOC_Os07g38240 | 非生物胁迫 | 低温萌发能力降低 | 野生型 | 低温萌发能力提高 |
| OsTPP7 | LOC_Os09g20390 | 非生物胁迫 | 厌氧萌发耐受性增加 | 突变型 | 厌氧萌发耐受性增加 |
| HAN1 | LOC_Os11g29290 | 非生物胁迫 | 耐冷性消失 | 野生型 | 耐冷性强 |
| qLTG3-1 | LOC_Os03g01320 | 非生物胁迫 | 耐低温萌发能力提高 | 突变型 | 耐低温萌发能力提高 |
| GH3-2 | LOC_Os01g55940 | 生物胁迫 | 白叶枯病性性增强 | 野生型 | 白叶枯病性性一般 |
| Xa5 | LOC_Os05g01710 | 生物胁迫 | 白叶枯病抗性提高 | 野生型 | 白叶枯病抗性一般 |
| Xa23 | LOC_Os11g37620 | 生物胁迫 | 白叶枯病抗性增强 | 野生型 | 白叶枯病抗性一般 |
| Pib | LOC_Os02g57310 | 生物胁迫 | 稻瘟病抗性提高 | 野生型 | 稻瘟病抗性一般 |
| Bph3 | LOC_Os04g12580/ LOC_Os04g12540 | 生物胁迫 | 褐飞虱抗性提高 | 野生型 | 褐飞虱抗性降低 |
| BPH29 | LOC_Os06g01860 | 生物胁迫 | 褐飞虱抗性增强 | 突变型 | 褐飞虱抗性增强 |
| Bph9 | LOC_Os12g37290 | 生物胁迫 | 褐飞虱抗性提高 | 野生型 | 褐飞虱抗性一般 |
| STV11 | LOC_Os11g30910 | 生物胁迫 | 条纹病毒抗性增强 | 突变型 | 条纹病毒抗性增强 |
| Pi9 | LOC_Os06g17900 | 生物胁迫 | 稻瘟病广谱抗性增强 | 野生型 | 稻瘟病广谱抗性一般 |
| Pid3 | LOC_Os06g22460 | 生物胁迫 | 稻瘟病抗性增强 | 突变型 | 稻瘟病抗性增强 |
| Pigm | LOC_Os06g17900 | 生物胁迫 | 稻瘟病抗性强 | 野生型 | 稻瘟病抗性一般 |
| OsCERK1 | LOC_Os08g42580 | 生物胁迫 | 磷吸收效率提高、稻瘟病抗性增强 | 突变型 | 磷吸收效率提高、稻瘟病抗性增强 |
| Pi5-1 | LOC_Os09g15840 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pi56 | LOC_Os09g16000 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pi54 | LOC_Os11g42010 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pib | LOC_Os02g57310 | 生物胁迫 | 稻瘟病抗性提高 | 野生型 | 稻瘟病抗性一般 |
| Pi21 | LOC_Os04g32850 | 生物胁迫 | 稻瘟病抗性提高 | 野生型 | 稻瘟病抗性一般 |
| Pi2 | LOC_Os06g17900 | 生物胁迫 | 稻瘟病广谱抗性增强 | 野生型 | 稻瘟病广谱抗性一般 |
| RGA5 | LOC_Os11g11810 | 生物胁迫 | 稻瘟病抗性增强 | 突变型 | 稻瘟病抗性增强 |
| Pi54 | LOC_Os11g42010 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pik | LOC_Os11g42010 | 生物胁迫 | 稻瘟病抗性增强 | 突变型 | 稻瘟病抗性增强 |
| OsbHLH153 | LOC_Os03g07540 | 植株形态 | 剑叶角变大 | 野生型 | 剑叶角小 |
| TAC3 | LOC_Os10g32540 | 植株形态 | 分蘖角增大 | 野生型 | 分蘖角小 |
| SBI | LOC_Os05g35400 | 植株形态 | 株高增加 | 野生型 | 株高降低 |
| APO1 | LOC_Os06g45460 | 植株形态 | 抗倒伏、增加每穗粒数 | 突变型 | 抗倒伏、增加每穗粒数 |
| OsAAP6 | LOC_Os01g65670 | 口感品质 | 增加蛋白质含量 | 突变型 | 增加蛋白质含量 |
| chalk5 | LOC_Os05g06480 | 口感品质 | 垩白率降低 | 突变型 | 垩白率降低 |
| fgr | LOC_Os08g32870 | 口感品质 | 米粒有香味 | 野生型 | 米粒无香味 |
| OsNPF6.1 | LOC_Os01g01360 | 其他 | 氮肥利用效率提高 | 野生型 | 氮肥利用效率低 |
| SLB1 | LOC_Os01g50530/ LOC_Os01g50520 | 其他 | 分蘖增多 | 突变型 | 分蘖增多 |
| OsNR2 | LOC_Os02g53130 | 其他 | 氮肥利用效率提高 | 突变型 | 氮肥利用效率提高 |
| NRT1.1B | LOC_Os10g40600 | 其他 | 氮肥利用效率提高 | 突变型 | 氮肥利用效率提高 |
| TOND1 | LOC_Os12g43440 | 其他 | 耐低氮胁迫 | 突变型 | 耐低氮胁迫 |
| BOC1 | LOC_Os03g12820 | 其他 | 愈伤组织容易褐化 | 突变型 | 愈伤组织容易褐化 |
| qSE3 | LOC_Os03g37930 | 其他 | 耐盐性提高 | 野生型 | 耐盐性一般 |
| 基因名称 | 基因号 | 农艺性状 | 突变型与野生型相比的表型变化 | 7-163S类型 | 7-163S表型 |
|---|---|---|---|---|---|
| NOG1 | LOC_Os01g54860 | 产量组成相关 | 每穗粒数增加 | 突变型 | 每穗粒数增加 |
| LAX1 | LOC_Os01g61480 | 产量组成相关 | 抗旱性提高、产量提高 | 突变型 | 抗旱性提高、产量提高 |
| DPL1 | LOC_Os01g15448 | 产量组成相关 | 杂交不育 | 野生型 | 杂交可育 |
| tms5 | LOC_Os02g12290 | 产量组成相关 | 温敏不育 | 突变型 | 温敏不育 |
| GW2 | LOC_Os02g14720 | 产量组成相关 | 籽粒变大,柱头不外露 | 野生型 | 籽粒变小 |
| OsGRF4 | LOC_Os02g47280 | 产量组成相关 | 氮肥吸收效率提高 | 野生型 | 氮肥吸收效率一般 |
| GNP1 | LOC_Os03g63970 | 产量组成相关 | 每穗粒数增加,株高增高 | 突变型 | 每穗粒数增加,株高增高 |
| NAL1 | LOC_Os04g52479 | 产量组成相关 | 增加叶宽、提高产量 | 突变型 | 增加叶宽、提高产量 |
| GW5 | LOC_Os05g09520 | 产量组成相关 | 粒宽减小 | 野生型 | 粒宽增加 |
| S5 | LOC_Os06g11010 | 产量组成相关 | 广亲和 | 野生型 | 籼粳杂交不亲和 |
| GW8 | LOC_Os08g41940 | 产量组成相关 | 粒宽增加,粒长减少 | 突变型 | 粒宽增加,粒长减少 |
| Rf4 | LOC_Os10g35240 | 产量组成相关 | 恢复野败型不育系育性 | 突变型 | 恢复野败型不育系育性 |
| LOX-3 | LOC_Os03g49260 | 产量组成相关 | 稻米储藏不产生腐败味 | 野生型 | 稻米储藏产生腐败味 |
| SKC1 | LOC_Os01g20160 | 非生物胁迫 | 耐盐性提高 | 野生型 | 耐盐性降低 |
| CYP72A31 | LOC_Os01g41800 | 非生物胁迫 | 除草剂耐受性增强 | 野生型 | 除草剂耐受性低 |
| NRAT1 | LOC_Os02g03900 | 非生物胁迫 | 铝胁迫敏感 | 野生型 | 耐铝胁迫 |
| OsHMA4 | LOC_Os02g10290 | 非生物胁迫 | 铜含量降低 | 野生型 | 铜含量高 |
| CAL1 | LOC_Os02g41904 | 非生物胁迫 | 减少镉积累 | 野生型 | 镉积累 |
| qLTG3-1 | LOC_Os03g01320 | 非生物胁迫 | 耐低温萌发能力提高 | 突变型 | 耐低温萌发能力提高 |
| OsCd1 | LOC_Os03g02380 | 非生物胁迫 | 镉富集增多 | 突变型 | 镉富集增多 |
| OsJAZ1 | LOC_Os04g55920 | 非生物胁迫 | 降低根长、根重 | 野生型 | 提高根长、根重 |
| OsHKT2;1 | LOC_Os06g48810 | 非生物胁迫 | 钾利用效率降低 | 野生型 | 钾利用效率提高 |
| OsHMA3 | LOC_Os07g12900 | 非生物胁迫 | 富集镉 | 野生型 | 不富集镉 |
| OsNRAMP5 | LOC_Os07g15370 | 非生物胁迫 | 富集镉 | 突变型 | 富集镉 |
| OsSAP16 | LOC_Os07g38240 | 非生物胁迫 | 低温萌发能力降低 | 野生型 | 低温萌发能力提高 |
| OsTPP7 | LOC_Os09g20390 | 非生物胁迫 | 厌氧萌发耐受性增加 | 突变型 | 厌氧萌发耐受性增加 |
| HAN1 | LOC_Os11g29290 | 非生物胁迫 | 耐冷性消失 | 野生型 | 耐冷性强 |
| qLTG3-1 | LOC_Os03g01320 | 非生物胁迫 | 耐低温萌发能力提高 | 突变型 | 耐低温萌发能力提高 |
| GH3-2 | LOC_Os01g55940 | 生物胁迫 | 白叶枯病性性增强 | 野生型 | 白叶枯病性性一般 |
| Xa5 | LOC_Os05g01710 | 生物胁迫 | 白叶枯病抗性提高 | 野生型 | 白叶枯病抗性一般 |
| Xa23 | LOC_Os11g37620 | 生物胁迫 | 白叶枯病抗性增强 | 野生型 | 白叶枯病抗性一般 |
| Pib | LOC_Os02g57310 | 生物胁迫 | 稻瘟病抗性提高 | 野生型 | 稻瘟病抗性一般 |
| Bph3 | LOC_Os04g12580/ LOC_Os04g12540 | 生物胁迫 | 褐飞虱抗性提高 | 野生型 | 褐飞虱抗性降低 |
| BPH29 | LOC_Os06g01860 | 生物胁迫 | 褐飞虱抗性增强 | 突变型 | 褐飞虱抗性增强 |
| Bph9 | LOC_Os12g37290 | 生物胁迫 | 褐飞虱抗性提高 | 野生型 | 褐飞虱抗性一般 |
| STV11 | LOC_Os11g30910 | 生物胁迫 | 条纹病毒抗性增强 | 突变型 | 条纹病毒抗性增强 |
| Pi9 | LOC_Os06g17900 | 生物胁迫 | 稻瘟病广谱抗性增强 | 野生型 | 稻瘟病广谱抗性一般 |
| Pid3 | LOC_Os06g22460 | 生物胁迫 | 稻瘟病抗性增强 | 突变型 | 稻瘟病抗性增强 |
| Pigm | LOC_Os06g17900 | 生物胁迫 | 稻瘟病抗性强 | 野生型 | 稻瘟病抗性一般 |
| OsCERK1 | LOC_Os08g42580 | 生物胁迫 | 磷吸收效率提高、稻瘟病抗性增强 | 突变型 | 磷吸收效率提高、稻瘟病抗性增强 |
| Pi5-1 | LOC_Os09g15840 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pi56 | LOC_Os09g16000 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pi54 | LOC_Os11g42010 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pib | LOC_Os02g57310 | 生物胁迫 | 稻瘟病抗性提高 | 野生型 | 稻瘟病抗性一般 |
| Pi21 | LOC_Os04g32850 | 生物胁迫 | 稻瘟病抗性提高 | 野生型 | 稻瘟病抗性一般 |
| Pi2 | LOC_Os06g17900 | 生物胁迫 | 稻瘟病广谱抗性增强 | 野生型 | 稻瘟病广谱抗性一般 |
| RGA5 | LOC_Os11g11810 | 生物胁迫 | 稻瘟病抗性增强 | 突变型 | 稻瘟病抗性增强 |
| Pi54 | LOC_Os11g42010 | 生物胁迫 | 稻瘟病抗性增强 | 野生型 | 稻瘟病抗性一般 |
| Pik | LOC_Os11g42010 | 生物胁迫 | 稻瘟病抗性增强 | 突变型 | 稻瘟病抗性增强 |
| OsbHLH153 | LOC_Os03g07540 | 植株形态 | 剑叶角变大 | 野生型 | 剑叶角小 |
| TAC3 | LOC_Os10g32540 | 植株形态 | 分蘖角增大 | 野生型 | 分蘖角小 |
| SBI | LOC_Os05g35400 | 植株形态 | 株高增加 | 野生型 | 株高降低 |
| APO1 | LOC_Os06g45460 | 植株形态 | 抗倒伏、增加每穗粒数 | 突变型 | 抗倒伏、增加每穗粒数 |
| OsAAP6 | LOC_Os01g65670 | 口感品质 | 增加蛋白质含量 | 突变型 | 增加蛋白质含量 |
| chalk5 | LOC_Os05g06480 | 口感品质 | 垩白率降低 | 突变型 | 垩白率降低 |
| fgr | LOC_Os08g32870 | 口感品质 | 米粒有香味 | 野生型 | 米粒无香味 |
| OsNPF6.1 | LOC_Os01g01360 | 其他 | 氮肥利用效率提高 | 野生型 | 氮肥利用效率低 |
| SLB1 | LOC_Os01g50530/ LOC_Os01g50520 | 其他 | 分蘖增多 | 突变型 | 分蘖增多 |
| OsNR2 | LOC_Os02g53130 | 其他 | 氮肥利用效率提高 | 突变型 | 氮肥利用效率提高 |
| NRT1.1B | LOC_Os10g40600 | 其他 | 氮肥利用效率提高 | 突变型 | 氮肥利用效率提高 |
| TOND1 | LOC_Os12g43440 | 其他 | 耐低氮胁迫 | 突变型 | 耐低氮胁迫 |
| BOC1 | LOC_Os03g12820 | 其他 | 愈伤组织容易褐化 | 突变型 | 愈伤组织容易褐化 |
| qSE3 | LOC_Os03g37930 | 其他 | 耐盐性提高 | 野生型 | 耐盐性一般 |
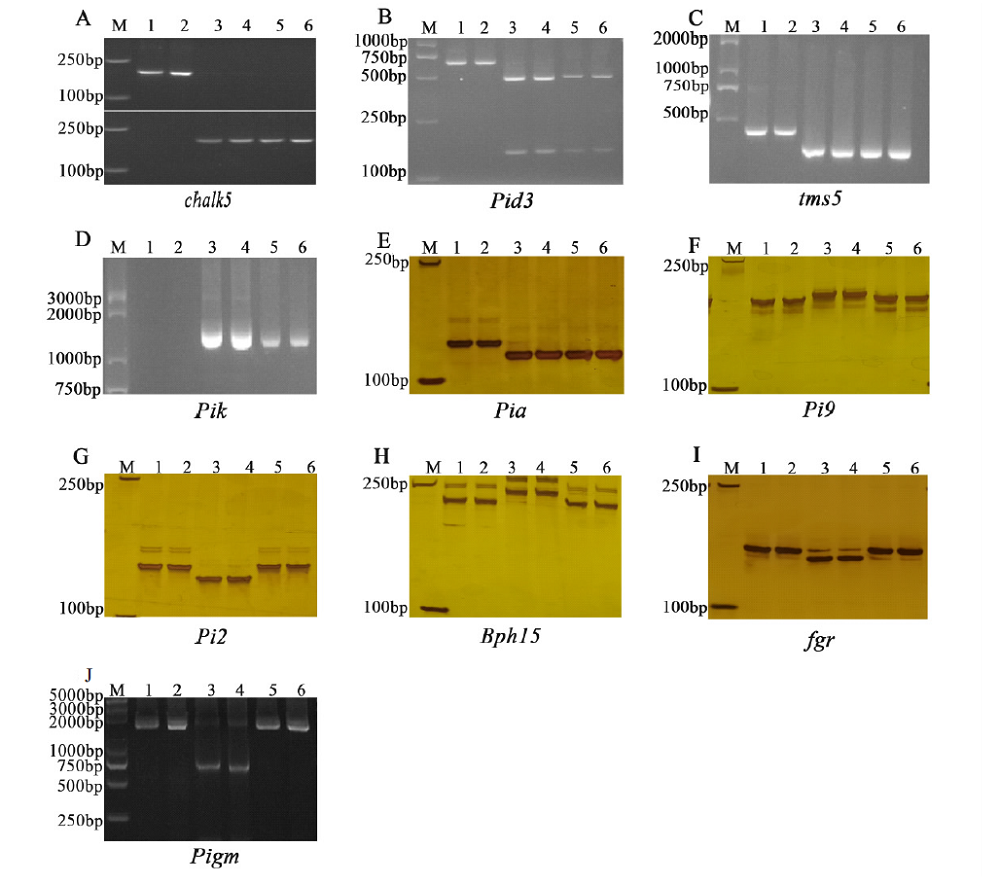
| [1] |
袁隆平. Progress in breeding of super hybrid rice[J]. 农学学报, 2018, 8(1):80-82.
doi: 10.11923/j.issn.2095-4050.cjas2018-1-080 |
| [2] |
袁隆平. 水稻的雄性不孕性[J]. 科学通报, 1966, 17(4):185-188.
|
| [3] |
胡兴明, 钱前. 我国杂交粳稻研究的回顾与思考[J]. 中国稻米, 2021, 27(4):9-11.
doi: 10.3969/j.issn.1006-8082.2021.04.003 |
| [4] |
石明松. 晚粳自然两用系选育及应用初报[J]. 湖北农业科学, 1981, 20(7):1-3.
|
| [5] |
符辰建, 胡小淳, 符星学, 等. 优质抗病高配合力中籼两用核不育系隆科638S的选育及应用[J]. 中国稻米, 2021, 27(3):61-66,69.
doi: 10.3969/j.issn.1006-8082.2021.03.013 |
| [6] |
白德朗, 罗孝和, 徐庆国. 两系超级杂交稻新组合培两优559的选育[J]. 湖南农业大学学报:自然科学版, 2002, 47(3):183-187.
|
| [7] |
杨联松, 白一松. 籼型优质光温敏核不育系1892S的选育[J]. 杂交水稻, 2006, 21(3):15-16,24.
|
| [8] |
杨振玉, 张国良, 张从合, 等. 中籼型优质光温敏核不育系广占63S的选育[J]. 杂交水稻, 2002, 17(4):8-10.
|
| [9] |
doi: 10.1038/s41588-020-00769-9 pmid: 33526925 |
| [10] |
郭韬, 余泓, 邱杰, 等. 中国水稻遗传学研究进展与分子设计育种[J]. 中国科学:生命科学, 2019, 49(10):1185-1212.
|
| [11] |
doi: 10.1016/j.cj.2021.05.001 URL |
| [12] |
姚姝, 陈涛, 骆名瑞, 等. 分子标记辅助选择定向改良武运粳7号的条纹叶枯病抗性[J]. 华北农学报, 2013, 28(4):195-203.
doi: 10.3969/j.issn.1000-7091.2013.04.036 |
| [13] |
田红刚, 陈红旗, 胡江, 等. 分子标记转育3个基因改良空育131稻瘟病抗性[J]. 核农学报, 2016, 30(11):2096-2103.
doi: 10.11869/j.issn.100-8551.2016.11.2096 |
| [14] |
刘维, 廖耀平, 卢东柏, 等. 分子标记技术聚合Wx基因改良水稻早熟不育系品质[J]. 分子植物育种, 2022, 20(14):4691-4699.
|
| [15] |
doi: 10.1093/nsr/nww006 URL |
| [16] |
夏加发, 李泽福, 唐光勇. 水稻新品种中籼898及其应用技术[J]. 作物杂志, 2001, 17(2):30.
|
| [17] |
贺闽, 尹俊杰, 冯志明, 等. 水稻稻瘟病和纹枯病抗性鉴定方法[J]. 植物学报, 2020, 55(5):577-587.
|
| [18] |
张爱芳, 王春林, 陈雨, 等. 安徽省水稻品种对水稻白叶枯病的抗性及白叶枯病小种鉴定[J]. 植物保护, 2012, 38(3):139-142.
|
| [19] |
廖俊杰. 采用CTAB法快速提取植物DNA[J]. 天津农业科学, 1993, 20(3):26.
|
| [20] |
华南农业大学. 稻瘟病抗性基因Pia功能特异性分子标记及其方法与应用[P]. 中国专利,CN201210163881.X, 2012-10-03.
|
| [21] |
吴金红, 蒋江松, 陈惠兰, 等. 水稻稻瘟病抗性基因Pi-2(t)的精细定位[J]. 作物学报, 2002, 84(4):505-509.
|
| [22] |
殷得所, 夏明元, 李进波, 等. 抗稻瘟病基因Pi9的STS连锁标记开发及在分子标记辅助育种中的应用[J]. 中国水稻科学, 2011, 25(1):25-30.
doi: 10.3969/j.issn.1001-7216.2011.01.004 |
| [23] |
梁毅, 杨婷婷, 谭令辞, 等. 水稻广谱抗瘟基因Pigm紧密连锁分子标记开发及其育种应用[J]. 杂交水稻, 2013, 28(4):63-68,74.
|
| [24] |
王天生, 陈惠清, 谢旺有, 等. 利用功能性分子标记辅助选育香型杂交水稻不育系[J]. 分子植物育种, 2020, 18(23):7801-7807.
|
| [25] |
李刚, 袁彩勇, 曹奎荣, 等. 544份水稻种质稻瘟病抗性鉴定及抗性基因的分布研究[J]. 中国农业大学学报, 2018, 23(5):22-28.
|
| [26] |
doi: 10.1534/genetics.109.102871 URL |
| [27] |
朱金燕, 王晴晴, 王军, 等. 水稻垩白基因Chalk5功能标记的开发与应用[J]. 华北农学报, 2017, 32(1):1-8.
doi: 10.7668/hbnxb.2017.01.001 |
| [28] |
安徽省农业科学院水稻所. 一个与水稻温敏不育基因tms5紧密连锁标记的开发与应用[P]. 中国专利,CN201110326770.1, 2012.06.20.
|
| [29] |
李进波, 夏明元, 戚华雄, 等. 水稻抗褐飞虱基因Bph14和Bph15的分子标记辅助选择[J]. 中国农业科学, 2006, 39(10):2132-2137.
|
| [30] |
|
| [31] |
doi: 10.1016/j.molp.2014.10.010 pmid: 25616388 |
| [32] |
张海珊, 章东方, 严丹侃, 等. 稻瘟病抗性基因对安徽省稻瘟病菌种群抗性的影响[J]. 中国稻米, 2020, 26(1):50-53.
doi: 10.3969/j.issn.1006-8082.2020.01.011 |
| [33] |
李珊, 杜春梅. 稻瘟病菌与水稻互作研究进展[J]. 中国农学通报, 2020, 36(24):125-131.
doi: 10.11924/j.issn.1000-6850.casb20190800517 |
| [34] |
徐如梦, 李冬月, 刘秀丽, 等. 水稻白叶枯病发病过程及抗病育种新思路[J]. 浙江农业科学, 2022, 63(1):114-120,123.
|
| [35] |
doi: 10.1111/pbi.2005.3.issue-3 URL |
| [36] |
doi: 10.1038/ng.2923 |
| [37] |
doi: 10.1038/s41467-019-10544-y |
| [38] |
doi: 10.1038/srep00286 |
| [39] |
戴青云, 刘代欢, 王德新, 等. 硅对水稻生长的影响及其缓解镉毒害机理研究进展[J]. 中国农学通报, 2020, 36(5):86-92.
doi: 10.11924/j.issn.1000-6850.casb18100012 |
| [1] | 陈书健, 陈京都, 杨呈芹, 许美刚, 高剑波. 不同施氮量对机械水条播水稻产量及品质的影响[J]. 中国农学通报, 2023, 39(9): 1-6. |
| [2] | 赵首萍, 肖文丹, 陈德, 叶雪珠, 张棋, 伍少福, 胡静, 高娜, 黄淼杰. 基于土壤质量和稻米安全的稻田重金属钝化效果评估[J]. 中国农学通报, 2023, 39(8): 51-62. |
| [3] | 刘伟喜, 尹文锋, 李小娟, 肖友伦. 旱改水田水稻旱青立病的发生原因及防治措施研究[J]. 中国农学通报, 2023, 39(8): 85-89. |
| [4] | 田婷, 张青, 徐雯. 基于无人机多光谱影像的水稻冠层SPAD值预测研究[J]. 中国农学通报, 2023, 39(4): 149-153. |
| [5] | 肖本泽, 王资霖. 转EPSPS基因抗除草剂水稻恢复系的培育及育种评价[J]. 中国农学通报, 2023, 39(2): 8-15. |
| [6] | 白玛仁增, 顿玉多吉, 德例归吉, 德吉央宗, 益西多吉, 边巴次仁. 星-地结合对水稻高温热害监测模型的研究[J]. 中国农学通报, 2023, 39(1): 133-141. |
| [7] | 罗先富, 刘文强, 潘孝武, 董铮, 刘三雄, 刘利成, 阳标仁, 盛新年, 李小湘. 应用剩余杂合体衍生的近等基因系定位水稻株高QTL[J]. 中国农学通报, 2022, 38(9): 1-5. |
| [8] | 黄钰, 陈斌, 肖关丽. 云南哈尼族地方水稻‘月亮谷’对褐飞虱取食危害的生理反应[J]. 中国农学通报, 2022, 38(9): 123-129. |
| [9] | 李兴华, 王欢, 张盛, 蔡星星, 周强, 周楠. 氮肥用量与运筹方式对晚籼稻产量及花后干物质积累与转运的影响[J]. 中国农学通报, 2022, 38(9): 6-13. |
| [10] | 王一凡, 劳晓璨, 余丽萍, 叶海龙. 水稻‘甬优15’分期播种的气象条件适宜性试验研究[J]. 中国农学通报, 2022, 38(7): 106-109. |
| [11] | 李雪枫, 王坚, 叶晓园, 张秀婷, 王丽学. 苦瓜植株水浸提液对水稻种子萌发和秧苗生长的影响[J]. 中国农学通报, 2022, 38(6): 1-7. |
| [12] | 闫蕴韬, 何兮, 张海清, 贺记外. 水稻种子耐贮性研究进展[J]. 中国农学通报, 2022, 38(5): 1-8. |
| [13] | 翟彩娇, 张蛟, 崔士友, 陈澎军. 盐逆境对耐盐水稻穗部性状及产量构成因素的影响[J]. 中国农学通报, 2022, 38(4): 1-9. |
| [14] | 孙悦萍, 周芹, 徐利剑, 任红波, 马文琼, 张晓波, 金海涛, 陈国峰, 刘峰, 董见南. QuEChERS-高效液相色谱-串联质谱法测定水稻中氯虫苯甲酰胺和噻虫胺的残留[J]. 中国农学通报, 2022, 38(36): 126-131. |
| [15] | 李荣田, 时柳, 黄丽莹, 刘长华. 利用分子选择培育水稻‘吉粳88’(hd2/hd4)导入系[J]. 中国农学通报, 2022, 38(33): 1-9. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||