Welcome to Chinese Agricultural Science Bulletin,
Chinese Agricultural Science Bulletin ›› 2026, Vol. 42 ›› Issue (12): 186-198.doi: 10.11924/j.issn.1000-6850.casb2025-0588
Previous Articles Next Articles
YANG Hanjing( ), ZHAO Jun, LI Hang, ZHANG Shuai()
), ZHAO Jun, LI Hang, ZHANG Shuai()
Received:2025-07-14
Revised:2026-01-08
Online:2026-06-25
Published:2026-06-23
CLC Number:
YANG Hanjing, ZHAO Jun, LI Hang, ZHANG Shuai. Research Progress on Chemical Control Status and Insecticide Resistance of Key Aphid Species[J]. Chinese Agricultural Science Bulletin, 2026, 42(12): 186-198.
Add to citation manager EndNote|Ris|BibTeX
URL: https://www.casb.org.cn/EN/10.11924/j.issn.1000-6850.casb2025-0588
| 蚜虫种类 | 食性及寄主 | 分布 | 作物受害情况 |
|---|---|---|---|
| 豌豆蚜 Acyrthosiphon pisum | 多食性;寄主包括豌豆、 蚕豆和苜蓿等豆科作物 | 广泛分布于整个 北半球 | 刺吸寄主植物维管束液体,造成植物生长迟缓、叶片卷曲、枯萎。传播30种不同的病毒,在美国造成6000万美元损失,在中国造成10%~30%的产量损失[ |
| 花生蚜(豆蚜) Aphis craccivora | 多食性;200余种寄主, 为害花生、豇豆等 大约50种作物[ | 起源于欧洲,分布于世界 大部分地区 | 喜食豆类的幼嫩部分,常聚集于幼苗、嫩梢、花柄、花瓣及嫩荚上为害。非防治豇豆田损失超过50%。直接取食和传播植物病毒可造成花生48%的产量损失[ |
| 黑豆蚜 Aphis fabae | 多食性;350种寄主, 为害豇豆、荞麦、甜菜 和玉米等作物[ | 主要分布于南美洲、非洲 和北半球温带地区 | 刺吸寄主植物维管束液体,传播植物病毒病,造成植物叶片变黄,卷缩和开花延迟,可造成50%以上的产量损失[ |
| 棉蚜(瓜蚜) Aphis gossypii | 多食性;900多种寄主, 为害棉花、黄瓜、西葫芦、 辣椒、百合等100多种作物[ | 广泛分布于世界各地 | 直接取食和分泌蜜露对棉花生产造成危害,棉花因棉蚜减产可达30%~40%[ |
| 绣线菊蚜虫(苹果黄蚜) Aphis spiraecola | 多食性;寄主有65个属, 为害包括苹果、柑橘、桃、 梨、山楂、绣线菊等作物[ | 20世纪广泛传播,已 分布于世界大部分 地区 | 具有明显的趋嫩习性,常群集于果树嫩梢、叶片背面刺吸为害,被害叶片出现褪绿斑点并卷缩,被害枝条发育迟缓、扭曲,为害严重时造成树势衰弱,落花落果,提早落叶[ |
| 俄罗斯麦蚜 Diuraphis noxia | 多食性;主要寄主是 禾本科植物,为害小麦、 大麦和硬质小麦等作物 | 最初分布于俄罗斯、伊朗、 阿富汗和地中海沿岸国家, 20世纪70年代,迅速 扩散到非洲、亚洲、欧洲、 中东、南美洲和北美洲[ | 具有独特的危害方式,在取食时分泌致害物质破坏寄主表皮细胞、含有木质素的组织和叶绿体,导致寄主黄化和坏死,新叶不能正常展开,受害部位普遍失绿。在极端情况下导致寄主植物死亡。冬小麦从分蘖期到开花期,每1%分蘖受侵染导致产量损失约0.5%[ |
| 萝卜蚜(菜缢管蚜) Lipaphis pseudobrassicae (1975—2000年被认为是Lipaphis erysimi)[ | 寡食性;为害白菜、芥菜、 油菜等十字花科植物 | 广泛分布于世界各地 | 群集取食十字花科植物嫩梢、嫩叶汁液,造成幼叶向下畸形卷缩,植株畸形、矮小,影响包心或结球,造成减产,严重时造成作物萎蔫甚至死亡,产量损失50%~100%[ |
| 马铃薯长管蚜 (大戟长管蚜) Macrosiphum euphorbiae | 多食性;20多科200多种 寄主,为害蔷薇科作物 及马铃薯 | 起源于北美洲,1917年 在欧洲土豆上发现,已 扩散至世界各地 | 常聚集于马铃薯植株上部取食,严重时造成植株上部叶片卷叶或出现坏死斑点,叶片从中间叶脉向上翻卷,叶缘坏死,最终导致植株死亡,类似病害症状。可传播40多种非持久性病毒和5种持久性病毒[ |
| 麦无网长管蚜 Metopolophium dirhodum | 多食性;几乎所有禾本科 植物都是其寄主。为害 小麦、燕麦、黑麦等 禾本科作物[ | 广泛分布于世界各地,是 英国、西班牙、德国等欧洲 国家和南美洲阿根廷等 国家小麦上的蚜虫优势种群 | 前期常分布于小麦倒2和倒3叶, 后期主要聚集在旗叶取食 |
| 桃蚜(烟蚜) Myzus persicae | 多食性;超过400种寄主, 为害桃、甘蓝、烟草、辣椒、 马铃薯等作物[ | 广泛分布于世界各地 | 以口针刺吸寄主植物幼嫩组织汁液,并排泄蜜露,诱发煤污病,可传播包括马铃薯卷叶病毒、马铃薯Y病毒在内的115种植物病毒[ |
| 玉米蚜 Rhopalosiphum maidis | 多食性;主要寄主为 禾本科植物,包括玉米、 高粱、小麦、谷子等多种作物 | 广泛分布于世界各地, 是温带和热带地区 最重要的谷物害虫之一 | 聚集于植物叶片、叶鞘和花序吸取汁液。取食为害会导致叶片斑驳变形、植株矮化,会损害花序导致植株不育。能够传播植物病毒和真菌病菌 |
| 禾谷缢管蚜 Rhopalosiphum padi | 多食性; 几乎所有禾本科植物都是其寄主,可以在其他单子叶植物和多种双子叶植物上建立种群 | 广泛分布于世界各地 | 前期常分布于小麦下部叶背面、叶鞘和根茎附近,后期可在麦穗为害。刺吸取食造成叶片发黄,麦苗生长停滞、分蘖减少,影响籽粒灌浆,造成籽粒不饱满、千粒重下降,引起减产。分泌蜜露和传播植物病毒也可造成产量损失[ |
| 麦二叉蚜 Schizaphis graminum | 多食性; 几乎所有禾本科植物均为其寄主,可以在其他单子叶植物和多种双子叶植物上建立种群 | 广泛分布于世界各地 | 麦二叉蚜畏光耐干旱,多分布在植株的中下部叶片取食,主要在小麦苗期造成危害 |
| 麦长管蚜 Sitobion avenae | 多食性; 寄主包括多种禾本科植物,主要危害小麦、大麦、燕麦,南方偶害水稻、玉米、甘蔗、荻草等 | 广泛分布于世界各地 | 是中国麦蚜中的优势种。小麦生长前期集中在叶片的正面或背面,抽穗后集中在穗上刺吸汁液,受害小麦生长缓慢,叶色变黄,分蘖减少,千粒重下降。传播大麦黄矮病毒,从而降低产量 |
| 苜蓿斑蚜 (三叶草彩斑蚜) Therioaphis trifolii | 多食性;以豆科植物 为寄主,主要为害苜蓿 | 原产于欧洲、地中海地区 和西南亚,目前至少在 80个国家有分布[ | 聚集于苜蓿的嫩茎、叶片、嫩芽和花器上吸取汁液。被害植株叶片卷缩弯曲,叶脉萎黄,蕾和花变黄脱落,生长发育停滞,植株矮化甚至枯萎。危害后的苜蓿营养成分流失,粗蛋白、粗脂肪含量降低,粗纤维含量增加,分泌的蜜露导致煤污病,降低干草质量[ |
| 蚜虫种类 | 食性及寄主 | 分布 | 作物受害情况 |
|---|---|---|---|
| 豌豆蚜 Acyrthosiphon pisum | 多食性;寄主包括豌豆、 蚕豆和苜蓿等豆科作物 | 广泛分布于整个 北半球 | 刺吸寄主植物维管束液体,造成植物生长迟缓、叶片卷曲、枯萎。传播30种不同的病毒,在美国造成6000万美元损失,在中国造成10%~30%的产量损失[ |
| 花生蚜(豆蚜) Aphis craccivora | 多食性;200余种寄主, 为害花生、豇豆等 大约50种作物[ | 起源于欧洲,分布于世界 大部分地区 | 喜食豆类的幼嫩部分,常聚集于幼苗、嫩梢、花柄、花瓣及嫩荚上为害。非防治豇豆田损失超过50%。直接取食和传播植物病毒可造成花生48%的产量损失[ |
| 黑豆蚜 Aphis fabae | 多食性;350种寄主, 为害豇豆、荞麦、甜菜 和玉米等作物[ | 主要分布于南美洲、非洲 和北半球温带地区 | 刺吸寄主植物维管束液体,传播植物病毒病,造成植物叶片变黄,卷缩和开花延迟,可造成50%以上的产量损失[ |
| 棉蚜(瓜蚜) Aphis gossypii | 多食性;900多种寄主, 为害棉花、黄瓜、西葫芦、 辣椒、百合等100多种作物[ | 广泛分布于世界各地 | 直接取食和分泌蜜露对棉花生产造成危害,棉花因棉蚜减产可达30%~40%[ |
| 绣线菊蚜虫(苹果黄蚜) Aphis spiraecola | 多食性;寄主有65个属, 为害包括苹果、柑橘、桃、 梨、山楂、绣线菊等作物[ | 20世纪广泛传播,已 分布于世界大部分 地区 | 具有明显的趋嫩习性,常群集于果树嫩梢、叶片背面刺吸为害,被害叶片出现褪绿斑点并卷缩,被害枝条发育迟缓、扭曲,为害严重时造成树势衰弱,落花落果,提早落叶[ |
| 俄罗斯麦蚜 Diuraphis noxia | 多食性;主要寄主是 禾本科植物,为害小麦、 大麦和硬质小麦等作物 | 最初分布于俄罗斯、伊朗、 阿富汗和地中海沿岸国家, 20世纪70年代,迅速 扩散到非洲、亚洲、欧洲、 中东、南美洲和北美洲[ | 具有独特的危害方式,在取食时分泌致害物质破坏寄主表皮细胞、含有木质素的组织和叶绿体,导致寄主黄化和坏死,新叶不能正常展开,受害部位普遍失绿。在极端情况下导致寄主植物死亡。冬小麦从分蘖期到开花期,每1%分蘖受侵染导致产量损失约0.5%[ |
| 萝卜蚜(菜缢管蚜) Lipaphis pseudobrassicae (1975—2000年被认为是Lipaphis erysimi)[ | 寡食性;为害白菜、芥菜、 油菜等十字花科植物 | 广泛分布于世界各地 | 群集取食十字花科植物嫩梢、嫩叶汁液,造成幼叶向下畸形卷缩,植株畸形、矮小,影响包心或结球,造成减产,严重时造成作物萎蔫甚至死亡,产量损失50%~100%[ |
| 马铃薯长管蚜 (大戟长管蚜) Macrosiphum euphorbiae | 多食性;20多科200多种 寄主,为害蔷薇科作物 及马铃薯 | 起源于北美洲,1917年 在欧洲土豆上发现,已 扩散至世界各地 | 常聚集于马铃薯植株上部取食,严重时造成植株上部叶片卷叶或出现坏死斑点,叶片从中间叶脉向上翻卷,叶缘坏死,最终导致植株死亡,类似病害症状。可传播40多种非持久性病毒和5种持久性病毒[ |
| 麦无网长管蚜 Metopolophium dirhodum | 多食性;几乎所有禾本科 植物都是其寄主。为害 小麦、燕麦、黑麦等 禾本科作物[ | 广泛分布于世界各地,是 英国、西班牙、德国等欧洲 国家和南美洲阿根廷等 国家小麦上的蚜虫优势种群 | 前期常分布于小麦倒2和倒3叶, 后期主要聚集在旗叶取食 |
| 桃蚜(烟蚜) Myzus persicae | 多食性;超过400种寄主, 为害桃、甘蓝、烟草、辣椒、 马铃薯等作物[ | 广泛分布于世界各地 | 以口针刺吸寄主植物幼嫩组织汁液,并排泄蜜露,诱发煤污病,可传播包括马铃薯卷叶病毒、马铃薯Y病毒在内的115种植物病毒[ |
| 玉米蚜 Rhopalosiphum maidis | 多食性;主要寄主为 禾本科植物,包括玉米、 高粱、小麦、谷子等多种作物 | 广泛分布于世界各地, 是温带和热带地区 最重要的谷物害虫之一 | 聚集于植物叶片、叶鞘和花序吸取汁液。取食为害会导致叶片斑驳变形、植株矮化,会损害花序导致植株不育。能够传播植物病毒和真菌病菌 |
| 禾谷缢管蚜 Rhopalosiphum padi | 多食性; 几乎所有禾本科植物都是其寄主,可以在其他单子叶植物和多种双子叶植物上建立种群 | 广泛分布于世界各地 | 前期常分布于小麦下部叶背面、叶鞘和根茎附近,后期可在麦穗为害。刺吸取食造成叶片发黄,麦苗生长停滞、分蘖减少,影响籽粒灌浆,造成籽粒不饱满、千粒重下降,引起减产。分泌蜜露和传播植物病毒也可造成产量损失[ |
| 麦二叉蚜 Schizaphis graminum | 多食性; 几乎所有禾本科植物均为其寄主,可以在其他单子叶植物和多种双子叶植物上建立种群 | 广泛分布于世界各地 | 麦二叉蚜畏光耐干旱,多分布在植株的中下部叶片取食,主要在小麦苗期造成危害 |
| 麦长管蚜 Sitobion avenae | 多食性; 寄主包括多种禾本科植物,主要危害小麦、大麦、燕麦,南方偶害水稻、玉米、甘蔗、荻草等 | 广泛分布于世界各地 | 是中国麦蚜中的优势种。小麦生长前期集中在叶片的正面或背面,抽穗后集中在穗上刺吸汁液,受害小麦生长缓慢,叶色变黄,分蘖减少,千粒重下降。传播大麦黄矮病毒,从而降低产量 |
| 苜蓿斑蚜 (三叶草彩斑蚜) Therioaphis trifolii | 多食性;以豆科植物 为寄主,主要为害苜蓿 | 原产于欧洲、地中海地区 和西南亚,目前至少在 80个国家有分布[ | 聚集于苜蓿的嫩茎、叶片、嫩芽和花器上吸取汁液。被害植株叶片卷缩弯曲,叶脉萎黄,蕾和花变黄脱落,生长发育停滞,植株矮化甚至枯萎。危害后的苜蓿营养成分流失,粗蛋白、粗脂肪含量降低,粗纤维含量增加,分泌的蜜露导致煤污病,降低干草质量[ |
| 农药名称 | 农药类型 | 作用靶标 | 防治对象 | 抗药性水平 |
|---|---|---|---|---|
| 吡虫啉 | 新烟碱类杀虫剂 | 烟碱型乙酰胆碱受体(nAChR)激动剂,干扰神经信号传导 | 麦二叉蚜 Schizaphis graminum | 室内麦二叉蚜种群对吡虫啉的耐受性水平,经致死中浓度吡虫啉连续胁迫麦二叉蚜10代后,麦二叉蚜对吡虫啉的敏感性显著降低,其LC50值为室内对照种群的5.29倍[ |
| 氟啶虫胺腈 | 亚砜亚胺类杀虫剂 | 同吡虫啉,乙酰胆碱受体(nAChR)激动剂 | 麦长管蚜 Sitobion avenae | 研究发现北京、河北、山西、江苏、山东、河南、湖北和陕西等地的小麦麦长管蚜种群对氟啶虫胺腈均处于中等至高水平抗性(抗性倍数12~328倍),表现为低敏状态[ |
| 高效氯氰菊酯 | 拟除虫菊酯类杀虫剂 | 钠离子通道调节剂 | 棉蚜(瓜蚜) Aphis gossypii | 分析华中地区的瓜蚜室内抗性品系交互抗性结果表明,瓜蚜室内抗性品系对高效氯氰菊酯产生中等水平交互抗性(RR=11.70倍)[ |
| 氰戊菊酯 | 拟除虫菊酯类杀虫剂 | 钠离子通道调节剂 | 大豆蚜 Aphis glycines | 分析测定交互抗性毒力结果显示,大豆蚜与氰戊菊酯产生中等水平交互抗性,抗性倍数为13.83倍[ |
| 苦参碱 | 植物源农药 | 毒杀作用与谷氨酸和γ-氨基丁酸系统的调节有关[ | 棉蚜(瓜蚜) Aphis gossypii | 分析测定0.5%苦参碱水剂对棉蚜药后1 d,防治效果为40.35%,药后7 d,防效为60.58%,药后14 d,0.5%苦参碱水剂的防效为85.06%[ |
| 毒死蜱 | 有机磷类杀虫剂 | 抑制乙酰胆碱酯酶(AChE)引起的神经毒性 | 麦长管蚜 Sitobion avenae | 测定毒死蜱对小麦蚜虫的防治效果发现药后1 d 45%毒死蜱对小麦蚜虫的防治效果达到94.98%,药后3 d的防效中,45%毒死蜱对小麦蚜虫的防治效果达到99.30%;药后7 d,防效达到99.32%[ |
| 抗蚜威 | 氨基甲酸酯类杀虫剂 | 乙酰胆碱酯酶抑制剂 | 棉蚜(瓜蚜) Aphis gossypii | 大多地区瓜蚜对抗蚜威产生了低至高等水平的抗性(RR=6.90~283倍),湖北荆州2个种群对抗蚜威产生了低等水平抗性,抗性倍数分别是6.90倍和7.43倍[ |
| 阿维菌素 | 微生物源杀虫剂 | 干扰神经递质γ-氨基丁酸(GABA) | 甘蓝蚜 Brevicoryne brassicae | 36%阿维菌素·吡蚜酮对甘蓝蚜虫的田间防效较理想,药后10 d防效均达90.43%以上,阿维菌素·吡蚜酮药后7、10 d,对甘蓝蚜的田间防效均高达88%,剂量越高防效越好[ |
| 农药名称 | 农药类型 | 作用靶标 | 防治对象 | 抗药性水平 |
|---|---|---|---|---|
| 吡虫啉 | 新烟碱类杀虫剂 | 烟碱型乙酰胆碱受体(nAChR)激动剂,干扰神经信号传导 | 麦二叉蚜 Schizaphis graminum | 室内麦二叉蚜种群对吡虫啉的耐受性水平,经致死中浓度吡虫啉连续胁迫麦二叉蚜10代后,麦二叉蚜对吡虫啉的敏感性显著降低,其LC50值为室内对照种群的5.29倍[ |
| 氟啶虫胺腈 | 亚砜亚胺类杀虫剂 | 同吡虫啉,乙酰胆碱受体(nAChR)激动剂 | 麦长管蚜 Sitobion avenae | 研究发现北京、河北、山西、江苏、山东、河南、湖北和陕西等地的小麦麦长管蚜种群对氟啶虫胺腈均处于中等至高水平抗性(抗性倍数12~328倍),表现为低敏状态[ |
| 高效氯氰菊酯 | 拟除虫菊酯类杀虫剂 | 钠离子通道调节剂 | 棉蚜(瓜蚜) Aphis gossypii | 分析华中地区的瓜蚜室内抗性品系交互抗性结果表明,瓜蚜室内抗性品系对高效氯氰菊酯产生中等水平交互抗性(RR=11.70倍)[ |
| 氰戊菊酯 | 拟除虫菊酯类杀虫剂 | 钠离子通道调节剂 | 大豆蚜 Aphis glycines | 分析测定交互抗性毒力结果显示,大豆蚜与氰戊菊酯产生中等水平交互抗性,抗性倍数为13.83倍[ |
| 苦参碱 | 植物源农药 | 毒杀作用与谷氨酸和γ-氨基丁酸系统的调节有关[ | 棉蚜(瓜蚜) Aphis gossypii | 分析测定0.5%苦参碱水剂对棉蚜药后1 d,防治效果为40.35%,药后7 d,防效为60.58%,药后14 d,0.5%苦参碱水剂的防效为85.06%[ |
| 毒死蜱 | 有机磷类杀虫剂 | 抑制乙酰胆碱酯酶(AChE)引起的神经毒性 | 麦长管蚜 Sitobion avenae | 测定毒死蜱对小麦蚜虫的防治效果发现药后1 d 45%毒死蜱对小麦蚜虫的防治效果达到94.98%,药后3 d的防效中,45%毒死蜱对小麦蚜虫的防治效果达到99.30%;药后7 d,防效达到99.32%[ |
| 抗蚜威 | 氨基甲酸酯类杀虫剂 | 乙酰胆碱酯酶抑制剂 | 棉蚜(瓜蚜) Aphis gossypii | 大多地区瓜蚜对抗蚜威产生了低至高等水平的抗性(RR=6.90~283倍),湖北荆州2个种群对抗蚜威产生了低等水平抗性,抗性倍数分别是6.90倍和7.43倍[ |
| 阿维菌素 | 微生物源杀虫剂 | 干扰神经递质γ-氨基丁酸(GABA) | 甘蓝蚜 Brevicoryne brassicae | 36%阿维菌素·吡蚜酮对甘蓝蚜虫的田间防效较理想,药后10 d防效均达90.43%以上,阿维菌素·吡蚜酮药后7、10 d,对甘蓝蚜的田间防效均高达88%,剂量越高防效越好[ |
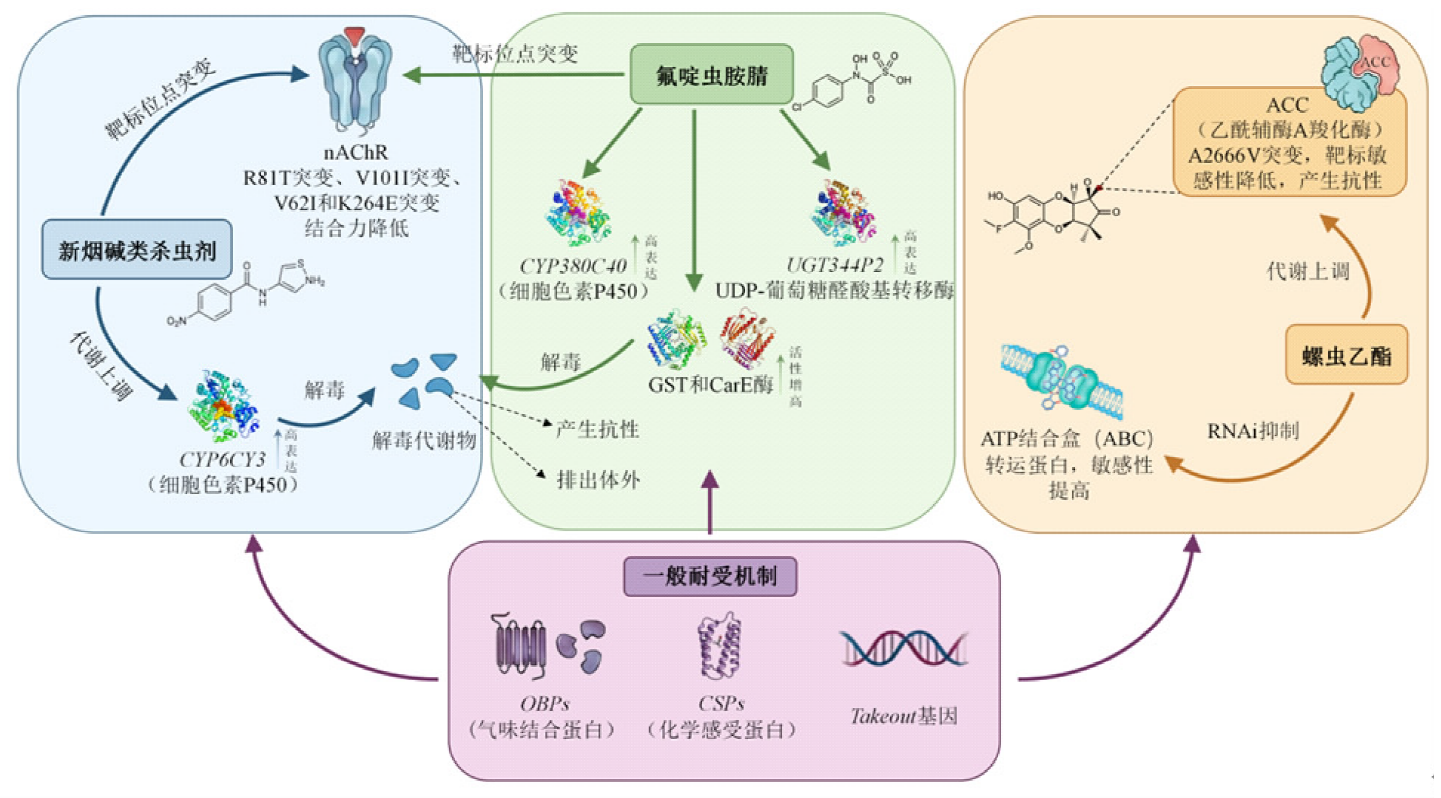
| 蚜虫种类 | 杀虫化合物 | 抗性机制 | 验证方法 |
|---|---|---|---|
| 桃蚜 Myzus persicae | 联苯菊酯 | VGSC突变(L1014F,M918T),转座子插入杂合子VGSC 等位基因编码序列导致提前终止[ | 生物信息学分析,分子生物学验证 |
| 高效氯氟氰菊酯 | 热激蛋白Hsp70高表达降低杀虫剂产生的氧胁迫[ | 在大肠杆菌中异源表达 | |
| 尼古丁、新烟碱类杀虫剂 | 启动子区域微卫星调控CYP6CY3的高表达[ 多拷贝复制、染色体重排导致CYP6CY3和CYP6CY4 表达增加,中肠和菌胞中高表达[ | 表达CYP6CY3的黑腹果蝇S2细胞[ | |
| 新烟碱类杀虫剂 | nAChR突变(R81T,V101I)[ | 抗性、敏感品系生物测定和 基因表达量分析[ | |
| 新烟碱类杀虫剂 | 躲避行为:蚜虫会主动从新烟碱类杀虫剂处理过的 叶片上迁飞到未处理过的叶片上[ | 行为学实验 | |
| 氟啶虫胺腈 | CYP380C40和UGT344P2表达量增加 (分别为21~76倍和6~33倍)[ | 转录组对比分析,转基因 黑腹果蝇和生物测定 | |
| 螺虫乙酯 | ACCase突变(A2666V)[ | 生物测定,分子检测 | |
| 棉蚜 Aphis gossypii | 拟除虫菊酯类 | VGSC突变(L1014F,M918L和M918V)[ | 生物测定,序列分析 |
| 溴氰虫酰胺、顺式氯氰菊酯 | P450(CYP380C6,CYP6CY21和CYP6CY7)和 UGTs(UGT341A4和UGT344M2)高表达[ | 转基因黑腹果蝇 | |
| 新烟碱类杀虫剂、溴氰虫酰胺 | 21个P450高表达[ | 外源表达,RNAi和转基因黑腹果蝇 | |
| 吡虫啉 | nAChR突变(R81T,V62I和K264E),nAChR基因表达量降低[ | 生物测定 | |
| 新烟碱类杀虫剂、螺虫乙酯、溴氰虫酰胺 | 化学感受蛋白AgoCSP4,AgosCSP5高表达[ | RNAi,转基因黑腹果蝇 | |
| 螺虫乙酯、新烟碱类 杀虫剂 | ABC转运蛋白和UGT高表达[ | 生物测定,RNAi和转基因黑腹果蝇 | |
| 螺虫乙酯 | ACCase14个氨基酸位点替换突变和过量表达[ | RNAi,酶活测定 | |
| 螺虫乙酯 | lncRNA调控CYP6CY21和CYP380C6的表达[ | RNAi,转基因黑腹果蝇 | |
| 氟啶虫胺腈 | P450活性增强(特别是CYP6CY13-2),CPR表达量变化[ | 酶活测定,RNAi | |
| 禾谷缢 管蚜Rhopalosiphum padi | 异丙威、氯氟氰菊酯 | 羧酸酯酶高表达[ | 体外表达,水解活性 |
| 拟除虫菊酯类 | VGSC突变(M918L)和P450代谢功能增强[ | 生物测定,序列分析和酶活测定 | |
| 高效氯氟氰菊酯、异丙威和吡虫啉 | P450和CPR高表达[ | RNAi、生物测定和基因 表达量分析 | |
| 拟除虫菊酯 | GPCR41高表达[ | RNAi | |
| 高效氯氰菊酯 | RpUGT344D38高表达[ | RNAi,体外表达和代谢活力检测 | |
| 吡虫啉、高效氯氰菊酯 | 化学感受蛋白和takeout基因高表达[ | RNAi | |
| 新烟碱类杀虫剂 | nAChR的突变和nAChR基因低表达(V53I,V53G, N54T,A60T,F61L,W79C和V83I)[ | 生物测定,序列分析 | |
| 麦长管蚜 Sitobion avenae | 高效氯氟氰菊酯 | VGSC突变(L1014F)[ | 突变品系生物测定 |
| 吡虫啉 | CYP6A14-1,CYP307A1,COE2和GST1-1-1表达量[ | RNAi(CYP6A14-1, CYP307A1,GST1-1-1和COE2) | |
| 荻草谷网蚜 Sitobion miscanthi | 吡虫啉 | miR-81调控SmUGGT1高表达,miR-3037调控 CYP6CY2高表达[ | RNAi,酶动力学和抑制试验 |
| 抗蚜威 | AChE1突变(S431F)和CYP6K1、CYP6A14高表达[ | RNAi | |
| 大豆蚜 Aphis glycines | 拟除虫菊酯 | VGSC突变(M918L,M918I,L925M,L1014F)[ | 序列分析,生物测定 |
| 吡虫啉 | CYP6CY7解毒代谢吡虫啉[ | RNAi,生物测定 | |
| 豌豆蚜 Acyrthosiphon pisum | 拟除虫菊酯 | CYP6CY12高表达[ | 表达量分析,异源表达 和代谢活性分析 |
| 黑豆蚜Aphis fabae | 毒死蜱 | 酯酶和ESTs酶活性升高[ | 酶活测定 |
| 蚜虫种类 | 杀虫化合物 | 抗性机制 | 验证方法 |
|---|---|---|---|
| 桃蚜 Myzus persicae | 联苯菊酯 | VGSC突变(L1014F,M918T),转座子插入杂合子VGSC 等位基因编码序列导致提前终止[ | 生物信息学分析,分子生物学验证 |
| 高效氯氟氰菊酯 | 热激蛋白Hsp70高表达降低杀虫剂产生的氧胁迫[ | 在大肠杆菌中异源表达 | |
| 尼古丁、新烟碱类杀虫剂 | 启动子区域微卫星调控CYP6CY3的高表达[ 多拷贝复制、染色体重排导致CYP6CY3和CYP6CY4 表达增加,中肠和菌胞中高表达[ | 表达CYP6CY3的黑腹果蝇S2细胞[ | |
| 新烟碱类杀虫剂 | nAChR突变(R81T,V101I)[ | 抗性、敏感品系生物测定和 基因表达量分析[ | |
| 新烟碱类杀虫剂 | 躲避行为:蚜虫会主动从新烟碱类杀虫剂处理过的 叶片上迁飞到未处理过的叶片上[ | 行为学实验 | |
| 氟啶虫胺腈 | CYP380C40和UGT344P2表达量增加 (分别为21~76倍和6~33倍)[ | 转录组对比分析,转基因 黑腹果蝇和生物测定 | |
| 螺虫乙酯 | ACCase突变(A2666V)[ | 生物测定,分子检测 | |
| 棉蚜 Aphis gossypii | 拟除虫菊酯类 | VGSC突变(L1014F,M918L和M918V)[ | 生物测定,序列分析 |
| 溴氰虫酰胺、顺式氯氰菊酯 | P450(CYP380C6,CYP6CY21和CYP6CY7)和 UGTs(UGT341A4和UGT344M2)高表达[ | 转基因黑腹果蝇 | |
| 新烟碱类杀虫剂、溴氰虫酰胺 | 21个P450高表达[ | 外源表达,RNAi和转基因黑腹果蝇 | |
| 吡虫啉 | nAChR突变(R81T,V62I和K264E),nAChR基因表达量降低[ | 生物测定 | |
| 新烟碱类杀虫剂、螺虫乙酯、溴氰虫酰胺 | 化学感受蛋白AgoCSP4,AgosCSP5高表达[ | RNAi,转基因黑腹果蝇 | |
| 螺虫乙酯、新烟碱类 杀虫剂 | ABC转运蛋白和UGT高表达[ | 生物测定,RNAi和转基因黑腹果蝇 | |
| 螺虫乙酯 | ACCase14个氨基酸位点替换突变和过量表达[ | RNAi,酶活测定 | |
| 螺虫乙酯 | lncRNA调控CYP6CY21和CYP380C6的表达[ | RNAi,转基因黑腹果蝇 | |
| 氟啶虫胺腈 | P450活性增强(特别是CYP6CY13-2),CPR表达量变化[ | 酶活测定,RNAi | |
| 禾谷缢 管蚜Rhopalosiphum padi | 异丙威、氯氟氰菊酯 | 羧酸酯酶高表达[ | 体外表达,水解活性 |
| 拟除虫菊酯类 | VGSC突变(M918L)和P450代谢功能增强[ | 生物测定,序列分析和酶活测定 | |
| 高效氯氟氰菊酯、异丙威和吡虫啉 | P450和CPR高表达[ | RNAi、生物测定和基因 表达量分析 | |
| 拟除虫菊酯 | GPCR41高表达[ | RNAi | |
| 高效氯氰菊酯 | RpUGT344D38高表达[ | RNAi,体外表达和代谢活力检测 | |
| 吡虫啉、高效氯氰菊酯 | 化学感受蛋白和takeout基因高表达[ | RNAi | |
| 新烟碱类杀虫剂 | nAChR的突变和nAChR基因低表达(V53I,V53G, N54T,A60T,F61L,W79C和V83I)[ | 生物测定,序列分析 | |
| 麦长管蚜 Sitobion avenae | 高效氯氟氰菊酯 | VGSC突变(L1014F)[ | 突变品系生物测定 |
| 吡虫啉 | CYP6A14-1,CYP307A1,COE2和GST1-1-1表达量[ | RNAi(CYP6A14-1, CYP307A1,GST1-1-1和COE2) | |
| 荻草谷网蚜 Sitobion miscanthi | 吡虫啉 | miR-81调控SmUGGT1高表达,miR-3037调控 CYP6CY2高表达[ | RNAi,酶动力学和抑制试验 |
| 抗蚜威 | AChE1突变(S431F)和CYP6K1、CYP6A14高表达[ | RNAi | |
| 大豆蚜 Aphis glycines | 拟除虫菊酯 | VGSC突变(M918L,M918I,L925M,L1014F)[ | 序列分析,生物测定 |
| 吡虫啉 | CYP6CY7解毒代谢吡虫啉[ | RNAi,生物测定 | |
| 豌豆蚜 Acyrthosiphon pisum | 拟除虫菊酯 | CYP6CY12高表达[ | 表达量分析,异源表达 和代谢活性分析 |
| 黑豆蚜Aphis fabae | 毒死蜱 | 酯酶和ESTs酶活性升高[ | 酶活测定 |
| [1] |
|
| [2] |
石丹丹, 张帅, 梁沛. 棉蚜抗药性现状及治理策略[J]. 植物保护, 2023, 49(5):270-278.
|
| [3] |
|
| [4] |
张帅, 李超侠, 康颖, 等. 细胞色素P450基因在棉蚜生物型间的分化[J]. 植物保护, 2023, 49(1):178-186.
|
| [5] |
doi: 10.1111/aen.v59.2 URL |
| [6] |
doi: 10.3390/plants13162332 URL |
| [7] |
|
| [8] |
王佩玲. 抗蚜棉在IPM体系中的作用及研究进展[J]. 中国棉花, 2005, 32(S1):89-90.
|
| [9] |
doi: 10.1603/0022-0493-94.2.439 URL |
| [10] |
殷万东, 仇贵生, 闫文涛, 等. 绣线菊蚜对苹果成熟/幼嫩叶片的选择性与适生性[J]. 应用生态学报, 2013, 24(7):2000-2006.
|
| [11] |
|
| [12] |
郭良珍, 刘绍友, 苏丽. 小麦禾谷缢管蚜的危害损失和防治指标研究[J]. 植物保护, 2000(6):12-14.
|
| [13] |
doi: 10.1038/s41597-023-02179-y |
| [14] |
doi: 10.1017/S0007485323000093 URL |
| [15] |
汤秋玲, 马康生, 高希武. 蔬菜蚜虫抗药性现状及抗性治理策略[J]. 植物保护, 2016, 42(6):11-20.
|
| [16] |
doi: 10.1016/j.ibmb.2023.103937 URL |
| [17] |
doi: 10.1002/ps.2016.72.issue-4 URL |
| [18] |
doi: 10.1002/ps.v63:8 URL |
| [19] |
doi: 10.1002/ps.v80.11 URL |
| [20] |
doi: 10.1016/j.pestbp.2016.11.004 URL |
| [21] |
doi: 10.1002/ps.v79.3 URL |
| [22] |
doi: 10.1002/ps.v78.4 URL |
| [23] |
doi: 10.1016/j.chemosphere.2020.128747 URL |
| [24] |
doi: 10.1002/ps.v80.2 URL |
| [25] |
doi: 10.1002/ps.v78.11 URL |
| [26] |
doi: 10.1038/s42003-021-02373-x |
| [27] |
doi: 10.1016/j.pestbp.2015.04.007 URL |
| [28] |
doi: 10.1016/j.ibmb.2022.103743 URL |
| [29] |
doi: 10.1016/j.cropro.2021.105834 URL |
| [30] |
doi: 10.1016/j.pestbp.2021.104981 URL |
| [31] |
doi: 10.1016/j.cropro.2022.105992 URL |
| [32] |
doi: 10.1016/j.cropro.2024.106783 URL |
| [33] |
doi: 10.1016/j.pestbp.2023.105504 URL |
| [34] |
doi: 10.1016/j.cropro.2019.105020 URL |
| [35] |
谢佳燕, 骆李涵. 吡虫啉多代胁迫对麦二叉蚜抗性发展的影响[J]. 武汉轻工大学学报, 2024, 43(5):63-68.
|
| [36] |
石丹丹. 华中地区瓜蚜抗药性监测及其对高效氯氟氰菊酯抗性机理研究[D]. 武汉: 华中农业大学, 2022.
|
| [37] |
朱斌, 梁沛. 害虫对杀虫剂抗性的发生与治理[J]. 现代农药, 2024, 23(4):1-6,37.
|
| [38] |
毕锐. 大豆蚜抗高效氯氟氰菊酯的分子机制及差异蛋白质组学分析[D]. 吉林: 吉林大学, 2016.
|
| [39] |
doi: 10.1002/ps.v76.12 URL |
| [40] |
胡迪, 张宣, 罗进仓, 等. 河西走廊地区棉蚜发生动态及植物源农药药效分析[J]. 新疆农业科学, 2019, 56(1):38-45.
doi: 10.6048/j.issn.1001-4330.2019.01.005 |
| [41] |
韦宁, 林璐璐, 何贤芳, 等. 8种药剂对小麦蚜虫的生物活性测定及田间药效试验[J]. 农药, 2020, 59(12):918-920,924.
|
| [42] |
王继英, 陈碧莲. 不同药剂对甘蓝蚜虫的田间药效试验[J]. 安徽农学通报, 2021, 27(3):67-68.
|
| [43] |
doi: 10.1002/ps.v33:2 URL |
| [44] |
doi: 10.1073/pnas.1314122110 URL |
| [45] |
doi: 10.1126/sciadv.aba1070 pmid: 32494722 |
| [46] |
doi: 10.3390/ijms20184521 URL |
| [47] |
doi: 10.1002/ps.v77.12 URL |
| [48] |
doi: 10.1584/jpestics.D17-006 URL |
| [49] |
doi: 10.3390/insects15030188 URL |
| [50] |
doi: 10.1021/acs.jafc.2c04867 URL |
| [51] |
doi: 10.1002/ps.v78.5 URL |
| [52] |
doi: 10.1016/j.pestbp.2024.105957 URL |
| [53] |
doi: 10.1002/ps.v76.8 URL |
| [54] |
doi: 10.1016/j.pestbp.2022.105088 URL |
| [55] |
doi: 10.1016/j.pestbp.2021.104879 URL |
| [56] |
doi: 10.1016/j.pestbp.2024.105958 URL |
| [57] |
doi: 10.1016/j.cbpb.2021.110682 URL |
| [58] |
doi: 10.1186/1471-2202-12-51 |
| [59] |
|
| [60] |
doi: 10.1371/journal.pgen.1009920 URL |
| [61] |
doi: 10.1002/ps.v78.4 URL |
| [62] |
doi: 10.1111/1744-7917.12116 pmid: 24677614 |
| [63] |
doi: 10.3390/insects12040335 URL |
| [64] |
doi: 10.3390/ijms23042356 URL |
| [65] |
doi: 10.1111/imb.v30.2 URL |
| [66] |
doi: 10.1016/j.pestbp.2020.104725 URL |
| [67] |
doi: 10.1111/mec.v29.14 URL |
| [68] |
doi: 10.1016/j.pestbp.2020.104582 URL |
| [69] |
doi: 10.1016/j.pestbp.2022.105187 URL |
| [70] |
doi: 10.1016/j.scitotenv.2023.162240 URL |
| [71] |
|
| [72] |
doi: 10.1016/j.cbpc.2023.109745 URL |
| [73] |
吕云彤. 棉蚜螺虫乙酯抗性ABC转运蛋白基因鉴定及其转录调控研究[D]. 吉林: 吉林大学, 2023.
|
| [74] |
doi: 10.1016/j.pestbp.2020.104583 URL |
| [75] |
doi: 10.1111/imb.2017.26.issue-4 URL |
| [76] |
doi: 10.1016/j.pestbp.2015.07.008 URL |
| [77] |
doi: 10.1021/acs.jafc.2c04263 URL |
| [78] |
doi: 10.1073/pnas.2100559118 URL |
| [79] |
doi: 10.1016/j.pestbp.2021.104995 URL |
| [80] |
doi: 10.1584/jpestics.D19-017 URL |
| [81] |
doi: 10.1002/ps.2013.70.issue-1 URL |
| [82] |
doi: 10.1111/jen.v145.3 URL |
| [83] |
doi: 10.1021/acs.jafc.1c00922 URL |
| [84] |
doi: 10.1016/j.pestbp.2022.105076 URL |
| [85] |
doi: 10.1016/j.pestbp.2020.104558 URL |
| [86] |
doi: 10.1021/acs.jafc.4c02682 URL |
| [87] |
doi: 10.3389/fphys.2018.00992 URL |
| [88] |
doi: 10.1371/journal.pone.0154633 URL |
| [89] |
doi: 10.1016/j.pestbp.2024.105894 URL |
| [90] |
doi: 10.1021/acs.jafc.3c08403 URL |
| [91] |
doi: 10.1021/acs.jafc.4c03368 URL |
| [92] |
doi: 10.1016/j.pestbp.2016.02.008 URL |
| [93] |
doi: 10.1002/ps.2014.70.issue-8 URL |
| [94] |
doi: 10.1016/j.cropro.2019.105014 URL |
| [95] |
doi: 10.1021/acs.jafc.4c02431 URL |
| [96] |
doi: 10.1038/s41598-022-16366-1 pmid: 35835854 |
| [97] |
doi: 10.1127/entomologia/2021/1226 URL |
| [98] |
doi: 10.1127/entomologia/2023/1848 URL |
| [99] |
doi: 10.1016/j.ecoenv.2020.111335 URL |
| [100] |
吴剑, 宋宝安. 绿色农药创新及靶标研究现状与思考[J]. 中国科学基金, 2020, 34(4):486-494.
|
| [101] |
doi: 10.1073/pnas.1913603117 URL |
| [102] |
|
| [103] |
doi: 10.1111/aen.12236 URL |
| [1] | WANG Jingwen, MENG Dewei, ZHOU Tianyue, CHEN Yongxin, ZHENG Linhao, HOU Xu, WU Yuwei, XU Wenjing. Research Status of Monolepta hieroglyphica [J]. Chinese Agricultural Science Bulletin, 2026, 42(9): 177-189. |
| [2] | LIU Hongmin, LIU Ruina, XIE Mingming, YANG Taiming. Research and Application of Spring Low Temperature Weather Index Insurance for Lu’an Crispy Peaches [J]. Chinese Agricultural Science Bulletin, 2026, 42(9): 124-129. |
| [3] | ZHAO Qianyun, LI Delu, WANG Fei, LI Ya. Response of Mucilaginous and Non-mucilage Artemisia ordosica Seed Germination to Salt Stress [J]. Chinese Agricultural Science Bulletin, 2026, 42(8): 60-66. |
| [4] | TAN Delong, CHENG Tanggen, QI Haijun, CAO Haishun, XIE Zhuolin, HUANG Xuhao, LIN Zeqin, WU Tingquan. Selection of Green Plant Protection Schemes for Leeks [J]. Chinese Agricultural Science Bulletin, 2026, 42(7): 176-183. |
| [5] | LIU Zhiwei, HUO Lizhi, SUN Longhua. Effects of Reduced Aphid Feeding on Survival and Oviposition of Coelophora biplagiata [J]. Chinese Agricultural Science Bulletin, 2026, 42(6): 201-209. |
| [6] | HUANG Wei, PAN Huixiao, WU Xuanke, HE Yan, TAN Mengxiang, LIU Yongyu. Analysis of Risk Zoning and Meteorological Factors of Double Cropping Rice Yield Disaster in Guangxi [J]. Chinese Agricultural Science Bulletin, 2026, 42(3): 144-154. |
| [7] | YANG Minfeng, MA Peng, ZENG Maolan, LUO Wanlin, WANG Yong, LIU Dongyang, YU Jiamin, GUO Shiping, WU Xinglong, PU Deqiang. Effects of Juvenile Hormone and Testosterone on Oviposition Ability of Coccinella septempunctata [J]. Chinese Agricultural Science Bulletin, 2026, 42(10): 139-143. |
| [8] | QIN Jiao, YAO Dandan, PEI Shuaishuai, GUO Shuhong, FENG Zhiyong, QIN Meng, TIAN Hongling. Advances in Common Chinese Zokor, Myospalax fontanieri (Rodentia: Spalacidae) in China [J]. Chinese Agricultural Science Bulletin, 2025, 41(8): 111-115. |
| [9] | LIANG Hui, DENG Quan, LIU Guo, CHEN Hezhu, MA Peng, LI Bin, LIU Dongyang, YU Jiamin, JIANG Lianqiang, PU Deqiang. Effect of Density on the Survival of Megalocaria dilatata in Different Stages [J]. Chinese Agricultural Science Bulletin, 2025, 41(8): 105-110. |
| [10] | LI Qin, ZHANG Qiaohui, QIN Zhanghui, ZHANG Zhexian, YIN Hongqing. Research Progress on Root Knot Nematode Disease of Codonopsis pilosula [J]. Chinese Agricultural Science Bulletin, 2025, 41(29): 107-112. |
| [11] | ZHAO Pengcheng, WANG Beibei, SUN Shuoyang, LIN Shijie, GAO Yu. Risk Assessment of Low-temperature Frost Disasters on Prunus domestica in Shihezi of Xinjiang [J]. Chinese Agricultural Science Bulletin, 2025, 41(27): 118-125. |
| [12] | BAO Jingwen, NI Dajing, HU Anxia, QIU Lifang, WANG Meishuo. Distribution Pattern of Heat Damage and Its Impact on Yield of Single-Season Rice in Xuancheng [J]. Chinese Agricultural Science Bulletin, 2025, 41(25): 100-107. |
| [13] | LIU Yan, HAN Wei. Field Control Effect of Different Chemical Treatments on Potato Late Blight [J]. Chinese Agricultural Science Bulletin, 2025, 41(2): 117-122. |
| [14] | LI Xin, WU Ruonan, ZHANG Yongchun, WANG Zhen, CHEN Minmin, YANG Liuyan. Evaluation of Heat Tolerance of Lilium davidii var. unicolor and Lilium Asiatic Hybrids [J]. Chinese Agricultural Science Bulletin, 2025, 41(16): 51-61. |
| [15] | CHEN Hongzhou, ZHANG Haibo, ZHOU Chen, ZHU Youli, YANG Hongfu, XU Chao, GUO Xiaomeng, ZHU Feng. Wheat Grain Deoxynivalenol (DON) Accumulation Dynamics and the Correlation Analysis of Its Influencing Factors [J]. Chinese Agricultural Science Bulletin, 2025, 41(15): 7-14. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||