欢迎访问《中国农学通报》,
中国农学通报 ›› 2023, Vol. 39 ›› Issue (17): 113-123.doi: 10.11924/j.issn.1000-6850.casb2022-0530
华雪铭( ), 康鹏, 魏翔(), 陈晨, 潘韵超, 王潜潜
), 康鹏, 魏翔(), 陈晨, 潘韵超, 王潜潜
收稿日期:2022-06-28
修回日期:2022-11-08
出版日期:2023-06-15
发布日期:2023-06-12
通讯作者:
魏翔,男,1995年出生,江苏连云港人,硕士,研究方向:水产动物营养学。通信地址:201306 上海市浦东新区沪城环路999号,E-mail:作者简介:华雪铭,女,1974年出生,浙江慈溪人,教授,博士,研究方向:水产动物营养与饲料学。通信地址:201306 上海市浦东新区沪城环路999号,E-mail:xmhua@shou.edu.cn。
基金资助:
HUA Xueming(), KANG Peng, WEI Xiang(), CHEN Chen, PAN Yunchao, WANG Qianqian
Received:2022-06-28
Revised:2022-11-08
Online:2023-06-15
Published:2023-06-12
摘要:
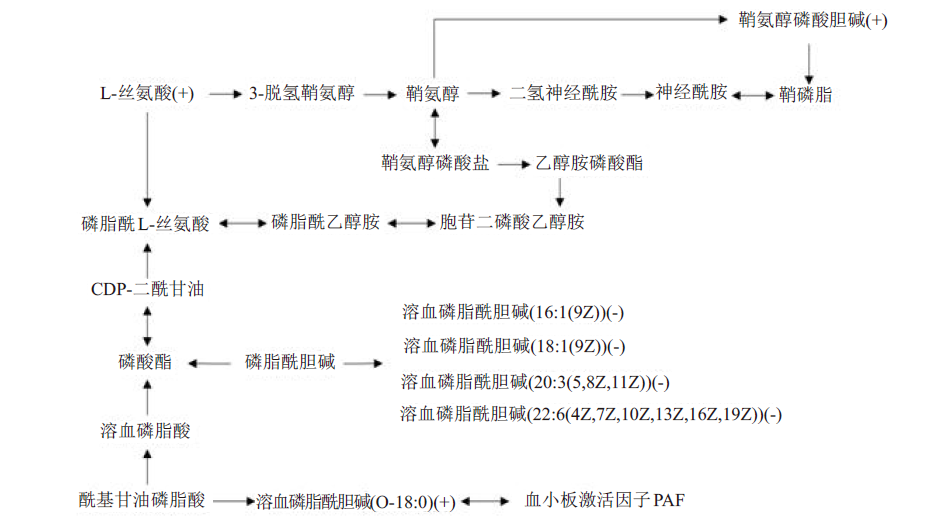
为了探究维生素K3在大口黑鲈体内的可能作用机制,在基础饲料中分别添加不同水平的维生素K3,配制成维生素K3含量分别为0.78(K30组)、15.84 mg/kg(K315组)的试验饲料,运用代谢组学和转录组学的方法对平均初始体重为(12.96±0.07) g、摄食饲料8周后生长和健康程度差异显著的K30组和K315组大口黑鲈肝脏进行分析。共鉴定出712个差异代谢物,以K30组为对照,K315组有326个上调的差异代谢物,386个下调;将差异代谢物注释到KEGG数据库中,共有13个差异代谢物,其中7个上调,6个下调。参与的功能通路有23条。整合通路表明维生素K3可通过L-丝氨酸调节鞘脂类代谢,也可通过壳寡糖调节糖类代谢。两组的差异表达基因在GO数据库中共有86个,参与的二级功能有26种。两组的差异基因在KEGG数据库中有29个,参与46条代谢途径。从肝脏代谢物和代谢通路的角度综合分析以上结果得到,大口黑鲈摄入适量的维生素K3可以提高抗氧化、免疫、机体恢复等能力,从而促进生长、维持健康。相关差异基因mRNA分析表明,维生素K3能够促进大口黑鲈骨骼发育和提高免疫能力。实验研究中没有发现与凝血有关的差异基因或代谢物。通过综合分析,在特定代谢组学和转录组学之间未发现相关性。因此,有必要进一步研究维生素K的作用机制。
华雪铭, 康鹏, 魏翔, 陈晨, 潘韵超, 王潜潜. 饲料维生素K3对大口黑鲈生长和健康的代谢转录调节机制研究[J]. 中国农学通报, 2023, 39(17): 113-123.
HUA Xueming, KANG Peng, WEI Xiang, CHEN Chen, PAN Yunchao, WANG Qianqian. Metabolic and Transcriptional Regulation of Dietary Vitamin K3 on Growth and Health of Largemouth Bass (Micropterus salmoides)[J]. Chinese Agricultural Science Bulletin, 2023, 39(17): 113-123.
| 原料/(g/kg干物质基础) | K30 |
|---|---|
| 鱼粉 | 450 |
| 豆粕 | 110 |
| 玉米蛋白粉 | 110 |
| 血粉 | 50 |
| α-淀粉 | 100 |
| 鱼油 | 40 |
| 菜籽油 | 20 |
| 大豆磷脂 | 30 |
| 多维(不含维生素K)* | 5 |
| 多矿* | 5 |
| 乌贼膏 | 10 |
| 氯化胆碱 | 5 |
| 磷酸二氢钙 | 10 |
| 牛磺酸 | 3 |
| 沸石粉 | 52 |
| 总计 | 1000 |
| 营养水平/% | |
| 粗蛋白 | 47.14 |
| 粗脂肪 | 12.16 |
| 总能** | 18.66 |
| 维生素K3 | 0.78 |
| 原料/(g/kg干物质基础) | K30 |
|---|---|
| 鱼粉 | 450 |
| 豆粕 | 110 |
| 玉米蛋白粉 | 110 |
| 血粉 | 50 |
| α-淀粉 | 100 |
| 鱼油 | 40 |
| 菜籽油 | 20 |
| 大豆磷脂 | 30 |
| 多维(不含维生素K)* | 5 |
| 多矿* | 5 |
| 乌贼膏 | 10 |
| 氯化胆碱 | 5 |
| 磷酸二氢钙 | 10 |
| 牛磺酸 | 3 |
| 沸石粉 | 52 |
| 总计 | 1000 |
| 营养水平/% | |
| 粗蛋白 | 47.14 |
| 粗脂肪 | 12.16 |
| 总能** | 18.66 |
| 维生素K3 | 0.78 |
| 基因 | 引物序列 | 产物大小 |
|---|---|---|
| CYP2R1 | F: TACGCCCTGCTCTACTTC R: CCTCCGTTTCTCTTCACT | 266 |
| nagB | F: CGGCTGGACTGAAGACCTTG R: GGCTGGGAGCAGACAGAATG | 237 |
| KL | F: ATGCCTGGCGGAATCTACTG R: GCTGATGTTGAAATGGGTGC | 111 |
| CPA1 | F: ACAGTGGTGGTCCCATTGCC R: AAGGAAGAGGTGCGAGTGAA | 280 |
| 基因 | 引物序列 | 产物大小 |
|---|---|---|
| CYP2R1 | F: TACGCCCTGCTCTACTTC R: CCTCCGTTTCTCTTCACT | 266 |
| nagB | F: CGGCTGGACTGAAGACCTTG R: GGCTGGGAGCAGACAGAATG | 237 |
| KL | F: ATGCCTGGCGGAATCTACTG R: GCTGATGTTGAAATGGGTGC | 111 |
| CPA1 | F: ACAGTGGTGGTCCCATTGCC R: AAGGAAGAGGTGCGAGTGAA | 280 |
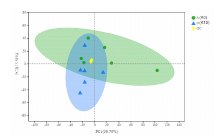






| K30A | K30B | K30C | K315A | K315B | K315C | |
|---|---|---|---|---|---|---|
| 原始序列数量 | 5.28×107 | 4.66×107 | 4.61×107 | 4.59×107 | 5.56×107 | 4.58×107 |
| 原始序列的碱基数量 | 7.98×109 | 7.04×109 | 6.96×109 | 6.94×109 | 8.39×109 | 6.92×109 |
| 质控序列数量 | 5.23×107 | 4.60×107 | 4.55×107 | 4.54×107 | 5.49×107 | 4.53×107 |
| 质控序列碱基数量 | 7.75×109 | 6.79×109 | 6.75×109 | 6.72×109 | 8.11×109 | 6.68×109 |
| 测序错误率 | 0.025 | 0.0266 | 0.026 | 0.0254 | 0.0254 | 0.0256 |
| Q20/% | 98.01 | 97.38 | 97.64 | 97.85 | 97.87 | 97.77 |
| Q30/% | 94.11 | 92.63 | 93.23 | 93.71 | 93.79 | 93.56 |
| GC content/% | 49.22 | 49.56 | 48.96 | 48.98 | 49.45 | 49.21 |
| K30A | K30B | K30C | K315A | K315B | K315C | |
|---|---|---|---|---|---|---|
| 原始序列数量 | 5.28×107 | 4.66×107 | 4.61×107 | 4.59×107 | 5.56×107 | 4.58×107 |
| 原始序列的碱基数量 | 7.98×109 | 7.04×109 | 6.96×109 | 6.94×109 | 8.39×109 | 6.92×109 |
| 质控序列数量 | 5.23×107 | 4.60×107 | 4.55×107 | 4.54×107 | 5.49×107 | 4.53×107 |
| 质控序列碱基数量 | 7.75×109 | 6.79×109 | 6.75×109 | 6.72×109 | 8.11×109 | 6.68×109 |
| 测序错误率 | 0.025 | 0.0266 | 0.026 | 0.0254 | 0.0254 | 0.0256 |
| Q20/% | 98.01 | 97.38 | 97.64 | 97.85 | 97.87 | 97.77 |
| Q30/% | 94.11 | 92.63 | 93.23 | 93.71 | 93.79 | 93.56 |
| GC content/% | 49.22 | 49.56 | 48.96 | 48.98 | 49.45 | 49.21 |
| K30A | K30B | K30C | K315A | K315B | K315C | |
|---|---|---|---|---|---|---|
| 质控序列数量 | 5.23×107 | 4.60×107 | 4.55×107 | 4.54×107 | 5.49×107 | 4.53×107 |
| 比对组装数量 | 3.81×107 | 3.41×107 | 3.33×107 | 3.31×107 | 3.95×107 | 3.31×107 |
| 比对组装率/% | 72.86 | 74.03 | 73.22 | 72.74 | 71.94 | 73.22 |
| K30A | K30B | K30C | K315A | K315B | K315C | |
|---|---|---|---|---|---|---|
| 质控序列数量 | 5.23×107 | 4.60×107 | 4.55×107 | 4.54×107 | 5.49×107 | 4.53×107 |
| 比对组装数量 | 3.81×107 | 3.41×107 | 3.33×107 | 3.31×107 | 3.95×107 | 3.31×107 |
| 比对组装率/% | 72.86 | 74.03 | 73.22 | 72.74 | 71.94 | 73.22 |


| 样本 | K30A | K30B | K30C | K315A | K315B | K315C |
|---|---|---|---|---|---|---|
| 平均值 | 24.89 | 23.96 | 21.90 | 16.52 | 20.81 | 22.47 |
| 最大值 | 19054.61 | 17782.79 | 18197.01 | 29512.09 | 39810.72 | 17378.01 |
| 上四分位数 | 13.49 | 13.80 | 14.45 | 12.88 | 13.49 | 13.80 |
| 中值 | 5.75 | 5.89 | 6.17 | 5.37 | 5.75 | 5.75 |
| 下四分位数 | 2.57 | 2.63 | 2.75 | 2.34 | 2.57 | 2.51 |
| 最小值 | 0.03 | 0.02 | 0.05 | 0.03 | 0.09 | 0.02 |
| 样本 | K30A | K30B | K30C | K315A | K315B | K315C |
|---|---|---|---|---|---|---|
| 平均值 | 24.89 | 23.96 | 21.90 | 16.52 | 20.81 | 22.47 |
| 最大值 | 19054.61 | 17782.79 | 18197.01 | 29512.09 | 39810.72 | 17378.01 |
| 上四分位数 | 13.49 | 13.80 | 14.45 | 12.88 | 13.49 | 13.80 |
| 中值 | 5.75 | 5.89 | 6.17 | 5.37 | 5.75 | 5.75 |
| 下四分位数 | 2.57 | 2.63 | 2.75 | 2.34 | 2.57 | 2.51 |
| 最小值 | 0.03 | 0.02 | 0.05 | 0.03 | 0.09 | 0.02 |

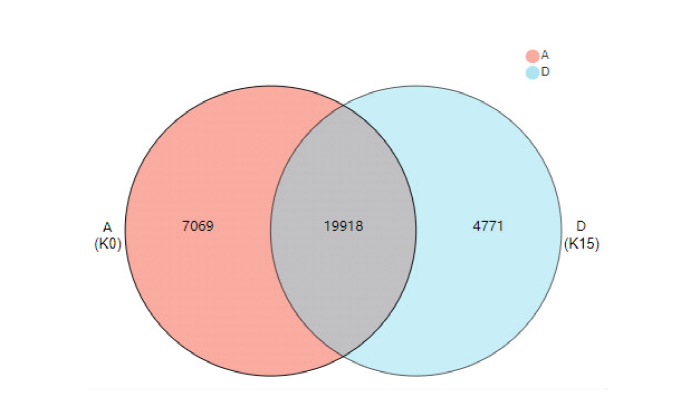

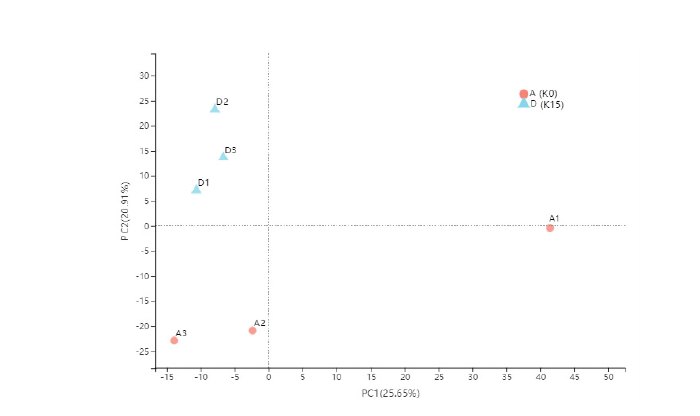
| 二级通路分类 | 通路名称 | KO | ||
|---|---|---|---|---|
| 生物降解和代谢 | 药物代谢-其他酶 | Ndk(-) | ||
| 碳水化合物代谢 | 戊糖、葡萄糖醛酸转换 | KL(+) | ||
| 氨基糖和核苷酸糖代谢 | E3.2.1.14(-);nagB (+) | |||
| 氨基酸代谢 | 酪氨酸代谢 | TYR(-) | ||
| 脂类代谢 | 类固醇生物合成 | CYP2R1 (+) | ||
| 初级胆汁酸的生物合成 | SCP2(-) | |||
| 不饱和脂肪酸的生物合成 | ACOT1_2_4(-) | |||
| 核苷酸代谢 | 嘌呤代谢 | ndk(-) | ||
| 嘧啶代谢 | ndk(-) | |||
| 信号转导 | FoxO信号通路 | BCL6(-) | ||
| Rap1信号通路 | THBS1(+) | |||
| NF-kappa B信号通路 | CCL21(-)、TNFSF11(+) | |||
| PI3K-Akt信号通路 | VWF(+) | |||
| TGF-beta信号通路 | THBS1(+) | |||
| 钙信号通路 | AGTR1(-) | |||
| Jak-STAT信号通路 | CISH(+) | |||
| 信号分子与相互作用 | 刺激神经组织中的相互作用 | NTSR1(-)、GRIA1(-) | ||
| cytokine-cytokine受体相互作用 | CCL21(-)、TNFSF11(+) | |||
| 细胞粘附分子(CAMs) | MHCⅡ(+)、CLDN(+)、LRRC4(+) | |||
| ECM-受体相互作用 | THBS1(+) | |||
| 运输和分解代谢 | 过氧化物酶体 | SCP2(-) | ||
| 吞噬体 | MHCⅡ(+)、THBS1(+) | |||
| 细胞群落-真核生物 | 紧密连接 | THBS1(+) | ||
| 局灶性粘连 | VWF(+) | |||
| 细胞生长与死亡 | p53信号通路 | THBS1(+) | ||
| 内分泌系统 | 甲状旁腺激素的合成、分泌和作用 | KL(+)、TNFSF11(+) | ||
| 黑素原生成 | TYR(-) | |||
| 肾素-血管紧张素系统 | MME(+) | |||
| PPAR信号通路 | SCP2(-) | |||
| 免疫系统 | 造血细胞谱系 | MHCⅡ(+)、MME(+) | ||
| Th17细胞分化 | MHCⅡ(+) | |||
| Th1和Th2细胞分化 | MHCⅡ(+) | |||
| 白细胞跨内皮转移 | CLDN(+) | |||
| 趋化因子信号通路 | CXCL1_2_3(-) | |||
| 产生IgA的肠道免疫网络 | MHCⅡ(+) | |||
| IL-17信号通路 | P38(-) | |||
| 抗原处理和呈递 | MHCⅡ(+) | |||
| 神经系统 | 多巴胺能突触 | ARNTL(-) | ||
| 谷氨酸能突触 | GRIK1(-) | |||
| 消化系统 | 蛋白质的消化和吸收 | CPA1(+) | ||
| 排泄系统 | 内分泌和其他因子调节的钙重吸收 | KL(+) | ||
| 二级通路分类 | 通路名称 | KO | ||
| 环境适应 | 昼夜夹带 | PER1(+) | ||
| 生理节律 | PER1(+)、NR1D1(+)、NR1F1(-)、ARNTL(-) | |||
| 生长 | 破骨细胞分化 | RANKL(+) | ||
| 轴突引导 | LRRC4(+) | |||
| 衰老 | 长寿调节途径 | KL(+) | ||
| 二级通路分类 | 通路名称 | KO | ||
|---|---|---|---|---|
| 生物降解和代谢 | 药物代谢-其他酶 | Ndk(-) | ||
| 碳水化合物代谢 | 戊糖、葡萄糖醛酸转换 | KL(+) | ||
| 氨基糖和核苷酸糖代谢 | E3.2.1.14(-);nagB (+) | |||
| 氨基酸代谢 | 酪氨酸代谢 | TYR(-) | ||
| 脂类代谢 | 类固醇生物合成 | CYP2R1 (+) | ||
| 初级胆汁酸的生物合成 | SCP2(-) | |||
| 不饱和脂肪酸的生物合成 | ACOT1_2_4(-) | |||
| 核苷酸代谢 | 嘌呤代谢 | ndk(-) | ||
| 嘧啶代谢 | ndk(-) | |||
| 信号转导 | FoxO信号通路 | BCL6(-) | ||
| Rap1信号通路 | THBS1(+) | |||
| NF-kappa B信号通路 | CCL21(-)、TNFSF11(+) | |||
| PI3K-Akt信号通路 | VWF(+) | |||
| TGF-beta信号通路 | THBS1(+) | |||
| 钙信号通路 | AGTR1(-) | |||
| Jak-STAT信号通路 | CISH(+) | |||
| 信号分子与相互作用 | 刺激神经组织中的相互作用 | NTSR1(-)、GRIA1(-) | ||
| cytokine-cytokine受体相互作用 | CCL21(-)、TNFSF11(+) | |||
| 细胞粘附分子(CAMs) | MHCⅡ(+)、CLDN(+)、LRRC4(+) | |||
| ECM-受体相互作用 | THBS1(+) | |||
| 运输和分解代谢 | 过氧化物酶体 | SCP2(-) | ||
| 吞噬体 | MHCⅡ(+)、THBS1(+) | |||
| 细胞群落-真核生物 | 紧密连接 | THBS1(+) | ||
| 局灶性粘连 | VWF(+) | |||
| 细胞生长与死亡 | p53信号通路 | THBS1(+) | ||
| 内分泌系统 | 甲状旁腺激素的合成、分泌和作用 | KL(+)、TNFSF11(+) | ||
| 黑素原生成 | TYR(-) | |||
| 肾素-血管紧张素系统 | MME(+) | |||
| PPAR信号通路 | SCP2(-) | |||
| 免疫系统 | 造血细胞谱系 | MHCⅡ(+)、MME(+) | ||
| Th17细胞分化 | MHCⅡ(+) | |||
| Th1和Th2细胞分化 | MHCⅡ(+) | |||
| 白细胞跨内皮转移 | CLDN(+) | |||
| 趋化因子信号通路 | CXCL1_2_3(-) | |||
| 产生IgA的肠道免疫网络 | MHCⅡ(+) | |||
| IL-17信号通路 | P38(-) | |||
| 抗原处理和呈递 | MHCⅡ(+) | |||
| 神经系统 | 多巴胺能突触 | ARNTL(-) | ||
| 谷氨酸能突触 | GRIK1(-) | |||
| 消化系统 | 蛋白质的消化和吸收 | CPA1(+) | ||
| 排泄系统 | 内分泌和其他因子调节的钙重吸收 | KL(+) | ||
| 二级通路分类 | 通路名称 | KO | ||
| 环境适应 | 昼夜夹带 | PER1(+) | ||
| 生理节律 | PER1(+)、NR1D1(+)、NR1F1(-)、ARNTL(-) | |||
| 生长 | 破骨细胞分化 | RANKL(+) | ||
| 轴突引导 | LRRC4(+) | |||
| 衰老 | 长寿调节途径 | KL(+) | ||


| [1] |
doi: 10.1146/nutr.2009.29.issue-1 URL |
| [2] |
|
| [3] |
doi: 10.1016/0305-0491(93)90304-N URL |
| [4] |
王洋, 徐奇友. 水产动物对维生素K需要量的研究进展[J]. 饲料研究, 2010(8):17-20.
|
| [5] |
许国旺, 路鑫, 杨胜利. 代谢组学研究进展[J]. 中国医学科学院学报, 2007, 29(6):701-711.
|
| [6] |
许彦阳, 姚桂晓, 刘平香, 等. 代谢组学在农产品营养品质检测分析中的应用[J]. 中国农业科学, 2019, 52(18):3163-3176.
doi: 10.3864/j.issn.0578-1752.2019.18.009 |
| [7] |
|
| [8] |
doi: 10.1038/227561a0 |
| [9] |
张强, 王悦, 李美秋, 等. 组学研究进展[J]. 河北医药, 2021, 43(11):1730-1734.
|
| [10] |
罗辉, 叶华, 肖世俊, 等. 转录组学技术在水产动物研究中的运用[J]. 水产学报, 2015, 39(4):598-607.
|
| [11] |
农业农村部渔业渔政管理局. 中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2022:33.
|
| [12] |
|
| [13] |
doi: 10.1111/j.1749-7345.2005.tb00139.x URL |
| [14] |
陈乃松, 马建忠, 周恒永, 等. 大口黑鲈对饲料中蛋氨酸需求量的评定[J]. 水产学报, 2010, 34(8):1244-1253.
|
| [15] |
|
| [16] |
doi: 10.1111/anu.2011.18.issue-1 URL |
| [17] |
doi: 10.1016/j.aquaculture.2014.10.013 URL |
| [18] |
doi: 10.1111/anu.12615 URL |
| [19] |
连雪原, 陈乃松, 王孟乐, 等. 大口黑鲈维生素A需求量[J]. 动物营养学报, 2017, 29(10):3819-3830.
|
| [20] |
doi: 10.1111/are.2011.43.issue-1 URL |
| [21] |
魏翔, 颜克涛, 戴宇峰, 等. 饲料中维生素K3含量对大口黑鲈消化能力、血清代谢指标、肌肉氨基酸和脂肪酸组成的影响[J]. 动物营养学报, 2022, 34(1):533-543.
doi: 10.3969/j.issn.1006-267x.2022.01.049 |
| [22] |
木晓云, 董跃伟, 温晓江, 等. 反相高效液相色谱法测定维生素K2软胶囊中K2(20)含量[J]. 分析试验室, 2009, 28(S1):33-35.
|
| [23] |
doi: 10.1016/j.aaf.2021.08.004 URL |
| [24] |
刘岩, 王慧, 史吉平, 等. 微生物法生产L-丝氨酸代谢工程研究进展[J]. 生物技术通报, 2015(8):44-49.
doi: 10.13560/j.cnki.biotech.bull.1985.2015.08.007 |
| [25] |
doi: 10.1194/jlr.M800635-JLR200 pmid: 19369694 |
| [26] |
时磊, 李玲燕, 付明婧, 等. 神经鞘氨醇磷酸胆碱对人真皮成纤维细胞表达MMP-1,MMP-3和TIMP-1的影响[J]. 中国皮肤性病学杂志, 2017(2):136-139.
|
| [27] |
pmid: 7693362 |
| [28] |
pmid: 6732789 |
| [29] |
pmid: 6808993 |
| [30] |
张义, 张志浩, 张继平. 壳寡糖在水产动物上的研究与应用[J]. 湖北农业科学, 2020, 59(6):5-9.
|
| [31] |
doi: 10.1007/s10499-017-0112-7 URL |
| [32] |
doi: 10.1002/bab.1513 pmid: 27245476 |
| [33] |
doi: 10.1016/j.fsi.2012.02.019 URL |
| [34] |
doi: 10.1016/j.fsi.2017.08.035 URL |
| [35] |
doi: 10.1093/czoolo/55.6.416 URL |
| [36] |
杨明星, 胡森. 脱氢抗坏血酸能够减轻脂多糖对细胞线粒体的损伤[J]. 中国危重病急救医学, 2010(4):213.
|
| [37] |
doi: 10.1016/j.pediatrneurol.2019.06.009 URL |
| [38] |
聂鑫, 张萍, 程刚, 等. 亚叶酸钙解救培美曲塞所致重度骨髓抑制[J]. 药物不良反应杂志, 2019(4):305-306.
doi: 10.3760/cma.j.issn.1008-5734.2019.04.017 |
| [39] |
李蓓, 孙庆敏, 黄磊. 注射用亚叶酸钙与亚叶酸钠的临床应用及分析[J]. 现代医药卫生, 2015(A1):64-66.
|
| [40] |
pmid: 10977869 |
| [41] |
doi: 10.1093/ndt/gfl178 URL |
| [42] |
智信, 陈晓, 苏佳灿. 破骨细胞研究新进展[J]. 中国骨质疏松杂志, 2019(9):1327-1330.
|
| [43] |
doi: 10.1016/j.bbrc.2004.09.073 pmid: 15465040 |
| [44] |
doi: 10.1007/s00251-018-1095-x pmid: 30421030 |
| [45] |
pmid: 12543708 |
| [46] |
吴静, 闵柔, 邬敏辰, 等. 羧肽酶研究进展[J]. 食品与生物技术学报, 2012(8):793-801.
|
| [1] | 徐大玮, 毕玉琦, 解恒杰, 刘睿. 基于紫外-可见吸收光谱对食品中残留添加剂的检测和分析[J]. 中国农学通报, 2023, 39(8): 127-132. |
| [2] | 梁展图, 全林发, 梁盛曦, 陈炳旭, 马群, 姚琼. 基于CiteSpace的鳞翅目昆虫转录组学研究态势分析[J]. 中国农学通报, 2023, 39(8): 142-148. |
| [3] | 李媛媛, 德力格尔, 刘晶, 石凯. 单细胞转录组在昆虫学研究的应用[J]. 中国农学通报, 2023, 39(17): 79-85. |
| [4] | 陈柳宏, 赵春雷, 王希, 李彦丽, 丁广洲, 陈丽. 单细胞转录组测序技术及其在植物研究中的应用[J]. 中国农学通报, 2022, 38(3): 87-93. |
| [5] | 陈明月, 姜涛, 赵冬梅, 白莉, 张雪琦, 孟姣. 先进的组学技术在植物抗病研究中的应用[J]. 中国农学通报, 2022, 38(24): 86-91. |
| [6] | 宋立民, 姜巨峰, 王宇, 徐晓丽, 丁子元, 郝俊, 徐林通. 不同养殖模式下2个群体大口黑鲈肌肉营养成分比较[J]. 中国农学通报, 2022, 38(14): 118-123. |
| [7] | 宋小双, 遇文婧, 闵凯, 周琦, 邓勋, 刘艳红, 姜瑞凤. 立枯丝核菌诱导下深色有隔内生真菌A024代谢组学分析[J]. 中国农学通报, 2021, 37(8): 25-32. |
| [8] | 王东鹏, 叶诚, 廖小丽. 微流控芯片在农产品安全检测中的应用[J]. 中国农学通报, 2021, 37(36): 148-154. |
| [9] | 周凡, 彭建, 陈刘浦, 朱凝瑜, 姚高华, 贝亦江, 马文君, 丁雪燕. 基于室内循环水系统的大口黑鲈3个品系养殖性能评估[J]. 中国农学通报, 2021, 37(24): 159-164. |
| [10] | 姜武, 翁国杭, 陈家栋, 叶传盛, 姜鑫凯, 陶正明. 基于LC-MS代谢组学的红杆与绿杆型多花黄精化学成分比较研究[J]. 中国农学通报, 2021, 37(17): 32-38. |
| [11] | 路正禹, 王堽, 李任任, 崔汝菲, 耿贵. 基于组学技术探究甜菜耐盐机理研究进展[J]. 中国农学通报, 2021, 37(15): 92-98. |
| [12] | 孙茜, 徐圣君, 曾贤桂, 张焕祯, 白志辉. 动物青贮饲料添加剂的研究进展[J]. 中国农学通报, 2020, 36(27): 158-164. |
| [13] | 李海威, 李应佳, 容思华, 吴耿璇, 张锦华, 张廉政, 祝思琪, 黄晓亮, 黄冠庆. 桑叶粉对1~28日龄‘麒麟’肉鸡生长性能与组织器官发育的影响[J]. 中国农学通报, 2020, 36(11): 113-117. |
| [14] | 耿 贵,吕春华,於丽华,李任任,王宇光. 甜菜组学技术研究进展[J]. 中国农学通报, 2019, 35(12): 124-129. |
| [15] | 王应芬,李龙兴,张明均,吴佳海. 酶和乳酸菌对多年生黑麦草与玉米秸秆混合青贮发酵品质的影响[J]. 中国农学通报, 2017, 33(5): 107-111. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||