欢迎访问《中国农学通报》,
中国农学通报 ›› 2025, Vol. 41 ›› Issue (2): 140-148.doi: 10.11924/j.issn.1000-6850.casb2024-0102
董猛1,2( ), 宋大鹏2, 王鲲鹏2, 丁仕波2, 王莹莹1, 李海鹏1, 杨峰山1, 付海燕1()
), 宋大鹏2, 王鲲鹏2, 丁仕波2, 王莹莹1, 李海鹏1, 杨峰山1, 付海燕1()
收稿日期:2024-02-20
修回日期:2024-05-15
出版日期:2025-01-13
发布日期:2025-01-13
通讯作者:
作者简介:董猛,男,1997年出生,内蒙古赤峰人,研究生,研究方向:茶树微生物与植物之间互作。通信地址:150080 黑龙江哈尔滨市学府路74号 黑龙江大学生命科学学院,E-mail:2508016755@qq.com。
基金资助:
DONG Meng1,2(), SONG Dapeng2, WANG Kunpeng2, DING Shibo2, WANG Yingying1, LI Haipeng1, YANG Fengshan1, FU Haiyan1()
Received:2024-02-20
Revised:2024-05-15
Published:2025-01-13
Online:2025-01-13
摘要:
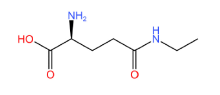
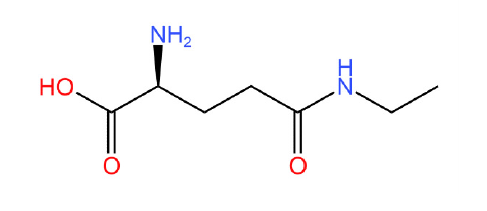
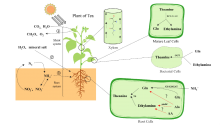
L-茶氨酸是茶叶中一种特殊的游离氨基酸,也是茶叶主要风味物质之一,因其具有放松心情、缓解疲劳、抗肿瘤、降血压等保健功能,被食品和医疗保健领域广泛关注。为了研究茶氨酸合成途径及方法,本文总结了目前L-茶氨酸合成的主要的途径,根据来源分为植物源合成和微生物源合成,根据合成方式分为生物酶法催化合成和化学方法合成,并分析了各个合成方法的优缺点,同时总结了L-茶氨酸在食品和医疗领域的应用,提出了高效合成茶氨酸未来的研究方向,以期为L-茶氨酸合成生产和应用提供参考。
董猛, 宋大鹏, 王鲲鹏, 丁仕波, 王莹莹, 李海鹏, 杨峰山, 付海燕. L-茶氨酸合成途径及应用展望[J]. 中国农学通报, 2025, 41(2): 140-148.
DONG Meng, SONG Dapeng, WANG Kunpeng, DING Shibo, WANG Yingying, LI Haipeng, YANG Fengshan, FU Haiyan. Synthesis Pathway and Application Prospect of L-theanine[J]. Chinese Agricultural Science Bulletin, 2025, 41(2): 140-148.

| 出版年份 | 菌株 | 关键基因 | 酶 | 底物和浓度 | |||
|---|---|---|---|---|---|---|---|
| 2013 | 硝基还原假单胞菌 | PnGLS | 重组谷氨酰胺酶 | L-谷氨酰胺0.3 mol/L、乙胺1.5 mol/L | |||
| 2013 | 大肠杆菌 | GGT | 谷氨酰胺转肽酶 | γ-谷氨酰苯胺0.1-10 mmol/L、乙胺2-200 mmol/L | |||
| 2014 | 地衣芽孢杆菌 | rBLGGT | 重组γ-谷氨酰胺转肽酶 | L-谷氨酰胺80 mmol/L、乙胺600 mmol/L | |||
| 2014 | 大肠杆菌 | GGT | γ-谷氨酰胺转肽酶 | L-谷氨酰胺200 mmol/L、乙胺2.2 mol/L | |||
| 2016 | 大肠杆菌 | GGT | 重组谷氨酰胺酶 | L-谷氨酰胺300 mmol/L、乙胺3 mol/L | |||
| 2016 | 大肠杆菌 | PPK/GMAS | 多聚磷盐激酶/γ-谷氨酰甲胺合成酶 | L-谷氨酸钠200 mmol/L、乙胺200 mmol/L | |||
| 2016 | 大肠杆菌 | GMAS | 多磷酸激酶 | L-谷氨酸钠200 mmol/L、乙胺200 mmol/L | |||
| 2017 | 木霉属 | GGT | 谷氨酰胺酶 | L-谷氨酰胺300 mmol/L、乙胺900 mmol/L | |||
| 2019 | 地衣芽孢杆菌 | BlGGT | γ-谷氨酰胺转肽酶 | L-谷氨酰胺250 mmol/L、乙胺600 mmol/L | |||
| 2019 | 硝基还原假单胞菌 | 谷氨酰酶 | L-谷氨酰胺0.3 mol/L、乙胺1 mol/L | ||||
| 2019 | Bacillus safensis | CseGGTs | 谷氨酰胺转肽酶 | L-谷氨酰胺20 mmol/L、乙胺20 mmol/L | |||
| 2019 | Bacillus subtlis | GGT | 重组谷氨酰胺酶 | L-谷氨酰胺0.25 mol/L、乙胺0.58 mol/L | |||
| 2019 | 大肠杆菌 | GMAS | γ-谷氨基甲酰胺合成酶 | 葡萄糖800 g/L、乙胺2 mol/L | |||
| 2020 | 大肠杆菌 | γ-GT | γ-谷氨酰胺转肽酶 | L-谷氨酰胺200 mmol/L、乙胺2 mol/L | |||
| 2022 | 枯草芽孢杆菌 | GGT | γ-谷氨酰转肽酶 | L-谷氨酰胺20 mmol/L、乙胺20 mmol/L | |||
| 2023 | 枯草芽孢杆菌 | BsGGT | γ-谷氨酰转移酶 | L-谷氨酰胺100 mmol/L、乙胺800 mmol/L | |||
| 2023 | 枯草芽孢杆菌 | GGT | γ-谷氨酰转肽酶 | L-谷氨酰胺200 mmol/L、乙胺1600 mmol/L | |||
| 出版年份 | 温度/℃ | pH | 时间/h | 转化率/% | 产率/% | 其他 | 参考文献 |
| 2013 | 37 | 10 | 5 | - | 40 | 反应添加0.3 U/mL GLS | [ |
| 2013 | 37 | 9 | 6 | 93 | - | 反应添加1 U/mL GGT | [ |
| 2014 | 37 | 9 | 3 | 87 | - | 反应添加0.5 U/mL GGT | [ |
| 2014 | 37 | 10 | 5 | 70 | [ | ||
| 2016 | 30 | 10.5 | 6 | 64.88 | - | 反应添加30 mg/mL湿菌体 | [ |
| 2016 | 37 | 7 | 24 | - | 86 | 反应添加5 mmol/LATP | [ |
| 2016 | 37 | 7 | 5 | - | 93 | 反应添加5 mmol/L ATP和无机多磷酸(polyp) | [ |
| 2017 | 37 | 8 | - | - | 43 | [ | |
| 2019 | 37 | 10.5 | 4 | 94 | - | 反应添加0.025 U/mL GGT | [ |
| 2019 | 30 | 9.5 | 11 | 66.1 | - | 反应添加15.5%蔗糖渗透冲击 | [ |
| 2019 | 28 | 7 | 98 | - | 90 | 内生菌发酵 | [ |
| 2019 | 30 | 10 | 16 | 74 | - | [ | |
| 2019 | 37 | 7 | 37 | - | 70 | [ | |
| 2020 | 55 | 10 | 5 | - | 83 | 反应添加0.02 U/mL BaGT | [ |
| 2022 | - | - | 87 | 45.2 | - | [ | |
| 2023 | 25 | 11.6 | 6 | - | 93 | [ | |
| 2023 | 35 | 10.5 | 12 | 95 | - | 反应添加2U/mL GGT | [ |
| 出版年份 | 菌株 | 关键基因 | 酶 | 底物和浓度 | |||
|---|---|---|---|---|---|---|---|
| 2013 | 硝基还原假单胞菌 | PnGLS | 重组谷氨酰胺酶 | L-谷氨酰胺0.3 mol/L、乙胺1.5 mol/L | |||
| 2013 | 大肠杆菌 | GGT | 谷氨酰胺转肽酶 | γ-谷氨酰苯胺0.1-10 mmol/L、乙胺2-200 mmol/L | |||
| 2014 | 地衣芽孢杆菌 | rBLGGT | 重组γ-谷氨酰胺转肽酶 | L-谷氨酰胺80 mmol/L、乙胺600 mmol/L | |||
| 2014 | 大肠杆菌 | GGT | γ-谷氨酰胺转肽酶 | L-谷氨酰胺200 mmol/L、乙胺2.2 mol/L | |||
| 2016 | 大肠杆菌 | GGT | 重组谷氨酰胺酶 | L-谷氨酰胺300 mmol/L、乙胺3 mol/L | |||
| 2016 | 大肠杆菌 | PPK/GMAS | 多聚磷盐激酶/γ-谷氨酰甲胺合成酶 | L-谷氨酸钠200 mmol/L、乙胺200 mmol/L | |||
| 2016 | 大肠杆菌 | GMAS | 多磷酸激酶 | L-谷氨酸钠200 mmol/L、乙胺200 mmol/L | |||
| 2017 | 木霉属 | GGT | 谷氨酰胺酶 | L-谷氨酰胺300 mmol/L、乙胺900 mmol/L | |||
| 2019 | 地衣芽孢杆菌 | BlGGT | γ-谷氨酰胺转肽酶 | L-谷氨酰胺250 mmol/L、乙胺600 mmol/L | |||
| 2019 | 硝基还原假单胞菌 | 谷氨酰酶 | L-谷氨酰胺0.3 mol/L、乙胺1 mol/L | ||||
| 2019 | Bacillus safensis | CseGGTs | 谷氨酰胺转肽酶 | L-谷氨酰胺20 mmol/L、乙胺20 mmol/L | |||
| 2019 | Bacillus subtlis | GGT | 重组谷氨酰胺酶 | L-谷氨酰胺0.25 mol/L、乙胺0.58 mol/L | |||
| 2019 | 大肠杆菌 | GMAS | γ-谷氨基甲酰胺合成酶 | 葡萄糖800 g/L、乙胺2 mol/L | |||
| 2020 | 大肠杆菌 | γ-GT | γ-谷氨酰胺转肽酶 | L-谷氨酰胺200 mmol/L、乙胺2 mol/L | |||
| 2022 | 枯草芽孢杆菌 | GGT | γ-谷氨酰转肽酶 | L-谷氨酰胺20 mmol/L、乙胺20 mmol/L | |||
| 2023 | 枯草芽孢杆菌 | BsGGT | γ-谷氨酰转移酶 | L-谷氨酰胺100 mmol/L、乙胺800 mmol/L | |||
| 2023 | 枯草芽孢杆菌 | GGT | γ-谷氨酰转肽酶 | L-谷氨酰胺200 mmol/L、乙胺1600 mmol/L | |||
| 出版年份 | 温度/℃ | pH | 时间/h | 转化率/% | 产率/% | 其他 | 参考文献 |
| 2013 | 37 | 10 | 5 | - | 40 | 反应添加0.3 U/mL GLS | [ |
| 2013 | 37 | 9 | 6 | 93 | - | 反应添加1 U/mL GGT | [ |
| 2014 | 37 | 9 | 3 | 87 | - | 反应添加0.5 U/mL GGT | [ |
| 2014 | 37 | 10 | 5 | 70 | [ | ||
| 2016 | 30 | 10.5 | 6 | 64.88 | - | 反应添加30 mg/mL湿菌体 | [ |
| 2016 | 37 | 7 | 24 | - | 86 | 反应添加5 mmol/LATP | [ |
| 2016 | 37 | 7 | 5 | - | 93 | 反应添加5 mmol/L ATP和无机多磷酸(polyp) | [ |
| 2017 | 37 | 8 | - | - | 43 | [ | |
| 2019 | 37 | 10.5 | 4 | 94 | - | 反应添加0.025 U/mL GGT | [ |
| 2019 | 30 | 9.5 | 11 | 66.1 | - | 反应添加15.5%蔗糖渗透冲击 | [ |
| 2019 | 28 | 7 | 98 | - | 90 | 内生菌发酵 | [ |
| 2019 | 30 | 10 | 16 | 74 | - | [ | |
| 2019 | 37 | 7 | 37 | - | 70 | [ | |
| 2020 | 55 | 10 | 5 | - | 83 | 反应添加0.02 U/mL BaGT | [ |
| 2022 | - | - | 87 | 45.2 | - | [ | |
| 2023 | 25 | 11.6 | 6 | - | 93 | [ | |
| 2023 | 35 | 10.5 | 12 | 95 | - | 反应添加2U/mL GGT | [ |
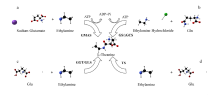

| 催化酶 | 简称 | 主要功能 | 催化反应条件 | 特点 | 参考文献 |
|---|---|---|---|---|---|
| γ-谷氨酰转肽酶 | GGT | 转运谷氨酰基 | 合成的pH值和 高浓度乙胺 | 反应不需要ATP、但需要高浓度的乙胺 抑制其水解反应 | [ |
| L-谷氨酰胺合成酶 | GS | 合成谷氨酸、谷氨酰胺 | 反应需要ATP和 较高的pH | 医学诊断中重要的参考指标、可催化植物 和合成氮素 | [ |
| γ-谷氨酰甲胺合成酶 | GMAS | 合成茶氨酸 | 需要ATP的参与 | 合成效率高 | [25]、[38]、[39] |
| γ-谷氨酰半胱合成酶 | γ-GCS | 合成合成γ-谷氨酰半胱氨酸 | 需要ATP的参与 | 可催化非氨基酸底物合成γ-谷氨酰半胱氨酸 | [ |
| L-谷氨酰胺酶 | GLS | 水解酰胺基 | 较高的底物浓度和pH | - | [28]、[42-43] |
| 茶氨酸合成酶 | TS | 合成茶氨酸 | 需要ATP的参与 | 只存在于茶科植物或小部分植物中, 不易分离纯化 | [ |
| 催化酶 | 简称 | 主要功能 | 催化反应条件 | 特点 | 参考文献 |
|---|---|---|---|---|---|
| γ-谷氨酰转肽酶 | GGT | 转运谷氨酰基 | 合成的pH值和 高浓度乙胺 | 反应不需要ATP、但需要高浓度的乙胺 抑制其水解反应 | [ |
| L-谷氨酰胺合成酶 | GS | 合成谷氨酸、谷氨酰胺 | 反应需要ATP和 较高的pH | 医学诊断中重要的参考指标、可催化植物 和合成氮素 | [ |
| γ-谷氨酰甲胺合成酶 | GMAS | 合成茶氨酸 | 需要ATP的参与 | 合成效率高 | [25]、[38]、[39] |
| γ-谷氨酰半胱合成酶 | γ-GCS | 合成合成γ-谷氨酰半胱氨酸 | 需要ATP的参与 | 可催化非氨基酸底物合成γ-谷氨酰半胱氨酸 | [ |
| L-谷氨酰胺酶 | GLS | 水解酰胺基 | 较高的底物浓度和pH | - | [28]、[42-43] |
| 茶氨酸合成酶 | TS | 合成茶氨酸 | 需要ATP的参与 | 只存在于茶科植物或小部分植物中, 不易分离纯化 | [ |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
doi: 10.1021/acs.jafc.7b02437 pmid: 28796499 |
| [7] |
李维, 向芬, 周凌云, 等. 氮素减施对茶树光合作用和氮肥利用率的影响[J]. 生态学杂志, 2020, 39(1):93-98.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
doi: 10.3389/fmicb.2015.01252 pmid: 26617590 |
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
张通, 龙科艺, 曹华杰, 等. 利用重组大肠杆菌发酵生产L-茶氨酸[J]. 食品与发酵工业, 2019, 45(22):6-11.
doi: 10.13995/j.cnki.11-1802/ts.021866 |
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
张飞, 熊吉滨, 刘均忠, 等. L-谷氨酰肼为底物酶法制备茶氨酸[J]. 精细化工, 2010, 27(2):130-132,41.
|
| [38] |
|
| [39] |
朱新雅. 利用γ-谷氨酰甲胺合成酶生物催化合成L-茶氨酸的研究[D]. 天津: 天津科技大学, 2019.
|
| [40] |
doi: 10.1007/s00253-020-10482-6 pmid: 32103318 |
| [41] |
|
| [42] |
孙帅. 谷氨酰胺酶和γ-谷氨酰转肽酶酶法合成L-茶氨酸的研究[D]. 南京: 南京大学, 2013.
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
潘鑫茹, 刘均忠, 张宏娟, 等. 共表达PPK和GMAS全细胞催化合成L-茶氨酸[J]. 精细化工, 2019, 36(9):1827-1832.
|
| [47] |
|
| [48] |
阎守和, MIGNOLET E, MEURENS M. 一种优化的茶氨酸化学合成方法简报[J]. 茶业通报, 2021, 43(4):154-155.
|
| [49] |
|
| [50] |
陈银霞, 李博. L-茶氨酸的性能及合成研究[J]. 化工技术与开发, 2011, 40(3):14-16.
|
| [51] |
国家卫生计生委. 关于批准茶叶茶氨酸为新食品原料等的公告(2014年第15号)[EB/OL]. http://www.hcbioworkx.com/uploadfile/202210/cfd9014fa180c98.pdf.
|
| [52] |
王锡洪, 梁慧玲, 毛斌瑀, 等. 茶氨酸的开发利用现状与展望[J]. 中国茶叶, 2021, 43(3):6-10.
|
| [53] |
帅玉英, 沈宇峰, 黄海娟, 等. 功能性配料L-茶氨酸的生产应用与法规情况简介[J]. 中国食品添加剂, 2013(2):181-185.
|
| [54] |
doi: 10.1021/acs.jafc.0c06694 pmid: 33475342 |
| [55] |
|
| [56] |
|
| [57] |
doi: 10.1016/j.jphs.2023.04.008 pmid: 37257943 |
| [58] |
刘洋, 李颂, 王春玲. 茶氨酸健康功效研究进展[J]. 食品研究与开发, 2016, 37(17):211-214.
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [1] | 翟玉凤, 丁兰, 余叶敏, 贾巧君, 梁宗锁, 汪得凯. 黄精皂苷的化学成分、生物合成及药理作用研究进展[J]. 中国农学通报, 2024, 40(29): 21-30. |
| [2] | 王莹, 李娟, 孙雪梅. 菊芋HtMYB2基因VIGS体系构建与功能验证[J]. 中国农学通报, 2024, 40(24): 116-121. |
| [3] | 罗秀芹, 韦卓文, 蔡杰, 安飞飞, 陈松笔, 薛晶晶. 转录组和代谢组联合分析阐释木薯叶片花青素合成机制[J]. 中国农学通报, 2024, 40(24): 100-106. |
| [4] | 马迪, 肖文芳, 李佐, 张俊卫, 陈和明, 吕复兵. 兰科植物花香成分研究进展[J]. 中国农学通报, 2023, 39(16): 52-60. |
| [5] | 杜明, 方玉, 李潜龙, 康云海, 王慧, 张从合. 玉米淀粉调控的研究进展[J]. 中国农学通报, 2021, 37(33): 8-14. |
| [6] | 万璐, 闫佳佳, 郑春英. 内生真菌发酵对刺五加异嗪皮啶合成的影响[J]. 中国农学通报, 2021, 37(13): 122-126. |
| [7] | 范雁婷, 樊若楠, 张辉, 李保珠. 植物黄酮醇生物合成及功能研究进展[J]. 中国农学通报, 2020, 36(33): 95-101. |
| [8] | 付金玉,刘 莉,杨旭辉,张乔玲,谢梅婷,华德平. 甜瓜苦味物质及其生物合成研究进展[J]. 中国农学通报, 2019, 35(8): 33-38. |
| [9] | 贾文斌,辛富刚,孙玉霞,姚川,丁汉凤. 隐甲藻生产二十二碳六烯酸的研究进展[J]. 中国农学通报, 2019, 35(30): 84-90. |
| [10] | 王春丽,王周礼,杨建利,苟升学,陈文杰. 大豆异黄酮的合成、调控、生理生态功能及其应用研究进展[J]. 中国农学通报, 2019, 35(19): 34-40. |
| [11] | 束红梅,郭书巧,巩元勇,蒋 璐,朱静雯,倪万潮. 西红花苷的生物合成及CCD家族基因研究进展[J]. 中国农学通报, 2016, 32(30): 91-95. |
| [12] | 赵晓菊 胡敏 尤凤丽 梁彦涛 邵小强. 吲哚乙酸生物合成及其结合物水解的研究进展[J]. 中国农学通报, 2014, 30(6): 254-259. |
| [13] | 张文 张树清 王学江. γ-聚谷氨酸的微生物合成及其在农业生产中的应用[J]. 中国农学通报, 2014, 30(6): 40-45. |
| [14] | 丁悦 黄科. 植物体内花青素累积的外源调控机制研究进展[J]. 中国农学通报, 2014, 30(19): 86-91. |
| [15] | 檀贝贝 孙 蕾 张克诚 崔增杰 杨振娟 武 哲. 抗生素生物合成基因簇的克隆策略[J]. 中国农学通报, 2011, 27(7): 206-209. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||